
Remember me







A female patient born as the first child to non-consanguineous parents of Turkish descent presented with recurrent fever episodes since the age of 12 months. Because the latter were associated with aphthous lesions and abdominal pain, she was clinically diagnosed with Familial Mediterranean Fever (FMF) despite no evidence of a disease-causing genetic variation in MEFV. However, the symptoms persisted under the treatment with colchicine. She was affected by IP, which was also present in her mother, and conical teeth (Fig. 1A, B). She had no history of severe infections and all vaccinations including BCG were well tolerated. Her family history was negative for infectious complications or autoinflammation.
Fig. 1
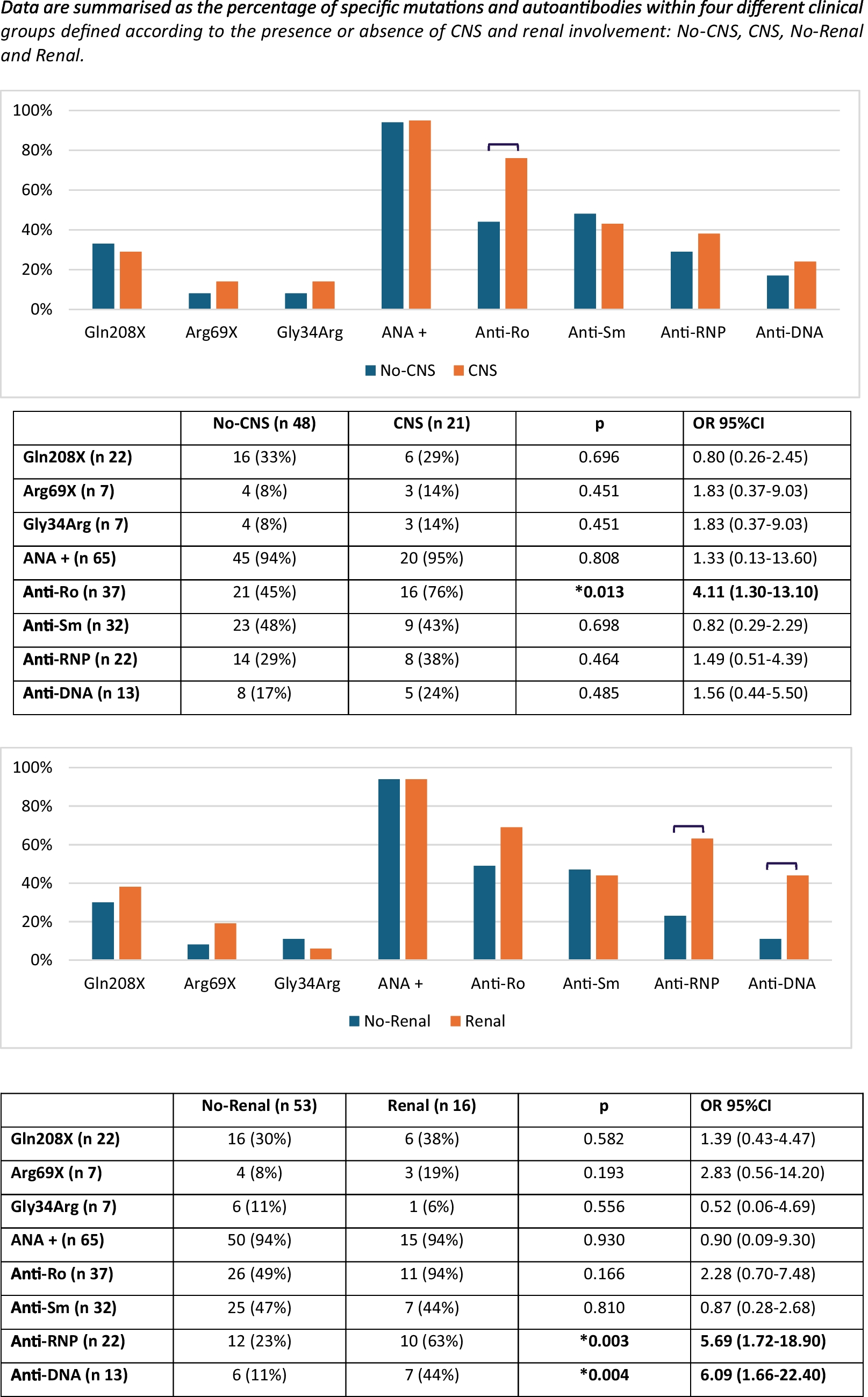
Clinical presentation of incontinentia pigmenti (IP) and NEMO-NDAS. At the age of 4y, the patient presented with ectodermal dysplasia characterized by hyperpigmentation along the Blashko lines (A) and conical teeth (B) (top: teeth at the age of 4y, bottom: dental X-ray at the age of 9y). Type I IFN-scores were calculated based on expression of IFN-stimulated genes analyzed in peripheral blood cells before start of treatment and under treatment with TNF inhibitors (C). IFN-scores of a cohort of 20 patients with typical interferonopathy (Aicardi-Goutières syndrome; AGS) are shown as control. Median values are displayed by horizontal bars. The cut-off for pathologic levels is indicated by the dotted line. Under treatment with TNF inhibitors as indicated, the patient developed mild hepatitis shown by elevated liver enzymes (ALT: alanine aminotransferase and AST: aspartate aminotransferase), and neutropenia (ANC: absolute neutrophil count) (D). Normal ranges are marked by grey-shaded areas
When she was investigated in our center at the age of 4 years, molecular genetics could not confirm fever syndromes caused by variations in MVK, NLRP3, or TNFRSF1A. Inflammation parameters such as IL-1β, TNF and IL-2R were in the normal range. Serum amyloid A was elevated during a flare (129 mg/l; normal range < 10 mg/l), but in normal ranges at later visits.
Immunophenotyping including immunoglobulin levels, and lymphocyte subsets revealed mildly impaired B-cell maturation with reduced CD27+ memory B cell subsets (Table S4). Vaccination titers for tetanus, diphtheria, and pneumococcal disease were non-protective (Table S4), however, the vaccination status was incomplete. The type I IFN-regulated gene signature was increased to similar ranges as observed in a historical cohort of patients with Aicardi-Goutières syndrome (AGS), which persisted after start of treatment with TNF inhibitors (Fig. 1C). After the diagnosis of autoinflammatory syndrome NEMO-NDAS, the patient started treatment with etanercept, which was later changed to adalimumab. However, diffuse alopecia with eczema leading to recurrent bacterial super-infections occurred under the latter medication. Although fever episodes and oral ulcers stopped, the patient developed neutropenia and elevated liver enzymes over time (Fig. 1D). Autoantibodies targeting liver tissue or neutrophils were not detected. A treatment attempt with the JAK inhibitor tofacitinib was not successful due to reoccurring fever episodes.
The p.Gln205* Variant Leads to Alternative Splicing and IKBKG-Δex5 TranscriptsBecause of the clinical presentation with ectodermal dysplasia a genetic variation in the X-chromosomal IKBKG gene was assumed and the heterozygous variant c.613 C > T p.(Gln205*) (NM_003639) in exon 5 was found in the patient and her mother. This variant has been reported before in a male patient with NEMO immunodeficiency syndrome and his family members [20]. Further characterization of this variant revealed expression of NEMO-Δex5 due to alternative splicing.
IKBKG transcripts were studied in peripheral blood cells obtained from the patient, her mother and 3 healthy controls. RNA was isolated from PBMCs, transcribed into cDNA and used for amplification of exons 3–8 (FigureS1A, B). Transcripts obtained by RT-PCR were separated by gel electrophoresis and identified by Sanger sequencing (Fig. 2A). Besides the full-length transcript variant 3 (NM_003639), several shorter transcripts were identified. The isoforms Δex4, Δex5, Δex4-5, and Δex4-6 were cloned into the pcDNA6 vector and used as size markers. Isoforms Dex4, and Dex4-5 result in disruption of the reading frame. Whereas the Δex4, Δex4-5, Δex4-6 isoforms could also be observed in the mother and the controls, Δex5 was predominantly found in patient cells.
Fig. 2
The variant c.613 C > T leads to expression of IKBKG-∆ex5 transcripts and NEMO-∆ex5 proteins. RNA was isolated from peripheral blood obtained from the patient, her mother and three healthy controls. IKBKG transcripts were amplified from cDNA and separated by electrophoresis (A). Full-length IKBKG transcripts (wt) and transcript variants ∆ex4 (∆4), ∆ex5 (∆5), ∆ex4-5 (∆4–5), ∆ex4-6 (∆4–6) were cloned into the pcDNA6 vector and used as size markers. In addition, bands obtained were excised and sequenced by Sanger sequencing: 1: wt (780 bp), 2: ∆4 (661 bp), 3 (*only observed in patient cells): ∆5 (627 bp), 4: ∆4–5 (508 bp), 5: ∆4–6 (411 bp). Transcript variants were further analyzed by nanopore sequencing and mapped gene reads were quantified using a library of all detected IKBKG transcripts. Shown are the frequencies of the most abundant variants ∆ex4, ∆ex5, ∆ex4-5, and ∆ex4-6 as % of all mapped gene reads obtained from whole PBMCs of patient, mother and controls (B). IKBKG transcripts were amplified from cDNA of T, B, NK lymphocytes, and monocytes and analyzed by nanopore sequencing (C-F). Shown are the frequencies of the most abundant variants Δex4, Δex5, Δex4-5, and Δex4-6 as % of all mapped gene reads. Of note, Δex4-5 could not be detected in B cells (D)
In addition, IKBKG transcripts were quantified using Oxford Nanopore Sequencing. Besides the full-length transcript, Δex4, Δex5, Δex4-5, and Δex4-6 transcripts were found in considerable amounts in all individuals (Fig. 2B), although Δex5 and Δex4-6 variants were increasingly expressed in patient PBMCs. Variants observed with lower frequency are summarized in the transcript table (Table S2).
Frequencies of IKBKG transcripts were further studied in isolated T, B, and NK lymphocytes, and monocytes (Fig. 2C-F, Figure S1C). In T cells, mostly Δex5 and Δex4-6 transcripts were found in the patient, whereas Δex5 was much rarer in her mother and the controls. The frequencies of WT and Δex5 transcripts were almost similar in NK cells, whereas only Δex4-6, or Δex5 could be observed in patient B cells and monocytes, respectively. In total, fewer reads were obtained from B cells and monocytes due to lower cell counts resulting in lower RNA content obtained from these subsets.
In summary, the distribution of IKBKG transcripts was shifted in patient blood cells to a higher frequency of Δex5 transcripts at the expense of the full-length transcript.
No Truncated Protein but NEMO-Δex5 is Expressed in Peripheral Blood CellsExpression of NEMO proteins were studied in lysed PBMCs obtained from the patient, her mother and controls using western blot. Proteins of respective IKBKG transcripts were synthesized in vitro to be used as size marker (Table S5). Two antibodies with different target sites, abNEMO150–300 binding to exon 4–7 (amino acids 150–300), and abNEMO278–396 binding to exon 7–10, respectively, were used for detection (Figure S1B). Expression of synthesized proteins was confirmed using the two antibodies mentioned above and GAPDH as loading control (Figure S1D). Besides the full-length NEMO protein (48 kDa), the truncated protein synthesized from the c.613 C > T variant could be detected by abNEMO150–300, whereas abNEMO278–396 detected NEMO-Δex5 and NEMO-Δex4-6, but not the truncated protein (Figure S1 E). As mentioned above, Δex4 and Δex4-5 transcripts do not allow protein expression due to disruption of the reading frame.
Analysis of NEMO expression in PBMC lysates revealed that no truncated protein was expressed by the patient or her mother (Fig. 3A). Whereas NEMO-∆ex4-6 protein could not be detected in any sample (Fig. 3B), NEMO-Δex5 was observed in patient lysates after prolonged separation up to the 37 kDa size marker (Fig. 3C). Full-length NEMO was expressed by all individuals tested.
Fig. 3
NEMO-∆ex5 is expressed in patient peripheral blood cells. Protein lysates obtained from PBMCs isolated from patient, mother and 3 controls were separated by SDS-PAGE and blotted on a PVDF membrane by western blotting. Proteins synthesized in-vitro from cloned full-length (wt), full-length with c.613 C > T variant in exon 5 (mut), ∆ex5 (∆5), and ∆ex4-6 (∆4–6) IKBKG transcripts were used as size markers in addition to a 10-250 kDa ladder. NEMO proteins were detected using primary antibodies binding to amino acids (aa) 150–300 (NEMO150–300) (A) and aa278-396 (NEMO278–396) (B, C).Besides the wt protein (48 kDa), several cross-reactional bands were detected by both antibodies, as described by the manufacturer. No protein could be detected from Δex4 and ∆ex4-5 transcripts due to disruption of the reading frame. The NEMO-∆ex5 protein could be observed in patient lysates following a longer period of separation (C). GAPDH was used as loading control, whereas bands resulting from non-specified proteins contained in the wheat germ extract were detected for in-vitro synthesized NEMO isoforms. Shown are representative western blots obtained from 3 experiments. In addition, NEMO expression was investigated in T, B, NK lymphocytes and monocytes by flow cytometry using abNEMO150–300 (left panel, blue histograms) and abNEMO278–396(right panel, red histograms) combined with anti-human CD3, CD19, CD56, and CD14 surface antibodies (D). Alexa fluor 488 anti-rabbit, and anti-mouse, respectively, were used as secondary antibodies and background controls (isotypes, grey histograms). In contrast to the controls, two peaks of intensity were observed in peripheral blood cell subsets obtained from the patient and her mother, which were quantified by gating
In summary, NEMO-∆ex5 expression could be detected in patient blood cells by abNEMO278–396, but not in cells obtained from her mother or controls.
NEMO and NEMO-Δex5 can be Discriminated as NEMObright and NEMOdim Populations in Peripheral Blood cell SubsetsNEMO expression was studied in T, B, and NK lymphocytes, and monocytes by flow cytometry using abNEMO278–396 and abNEMO150–300. Cell subsets were identified by surface markers gated as shown in Figure S2A. In contrast to healthy controls, two populations expressing NEMObright and NEMOdim could be detected by abNEMO278–396 in patient cells, and to a lower extend in her mother’s monocytes (Fig. 3D; Figure S2B). In contrast, only NEMObright expression could be found in cellular subsets of controls. The NEMOdim population could not be detected by abNEMO150–300. This investigation was performed 6 times leading to similar results, and twice in a different laboratory that included the analysis of granulocytes (Figure S2B). Interestingly, the distribution of NEMObright and NEMOdim populations among cellular subsets of the patient remained stable over time. We observed a NEMObright:NEMOdim ratio of approximately 75%:25% in T and B cells, an almost even (50%:50%) distribution in NK lymphocytes, and a ratio of 20%:80% in monocytes and granulocytes. A NEMObright:NEMOdim ratio of approximately 90%:10% was repeatedly found in the mother’s monocytes.
To compare the mean fluorescence intensities (MFIs) of full-length (WT) and Δex5 NEMO proteins, respectively, pcDNA6 vectors containing cDNAs of these variants were transfected into HCT116 colon carcinoma cells. Besides endogenous NEMO in non-transfected cells, cells transfected with WT, Δex5, or both variants were investigated for NEMO expression using abNEMO150–300 and abNEMO278–396 by flow cytometry. Consistent with results obtained by western blotting, NEMO-Δex5 was not detected by abNEMO150–300 but abNEMO278–396 (Figure S2C). However, differences between MFIs of NEMO-Δex5 and NEMO-WT proteins did not reach significance in three independent experiments (Figure S2D).
To investigate IKBKG variants transcribed in these populations, RNA was isolated from sorted NEMObright and NEMOdim populations obtained from patient PBMCs. Both full-length and ∆ex5 transcripts were amplified by RT-PCR (Figure S2E) as confirmed by Sanger sequencing. Since analyzed by end-point PCR, the number of transcripts could not be quantified.
Response to TNF-stimulation is Severely Diminished, Whereas Response to TLR Agonists is Mostly Retained in Peripheral Blood CellsNEMO function was investigated in whole blood obtained from the patient and her mother by quantification of IL-6 expression in response to stimulation with the TLR4-agonist LPS, the TLR2/6-agonist PAM2CSK4, IL-1β, and PMA/ionomycin (Fig. 4A), and IL-10 expression in response to TNF and PMA/ionomycin (Fig. 4B). Whereas the response to LPS was similar in patient and controls, the TNF response was diminished in patient cells compared to healthy controls. This assay was performed twice with similar results. The mother also showed slightly impaired responses upon stimulation with LPS and TNF as compared to reference values (Table S6).
Fig. 4
NEMO signaling function is partially impaired. Functional studies were performed on whole blood obtained from the patient and compared to shipping controls. Expression of IL-6 was measured in the supernatant following stimulation with LPS, PAM2CSK2, IL-1β and PMA/ionomycin (A). Levels of IL-10 were investigated after stimulation with TNF and PMA/ionomycin (B). Shown are the mean results with standard deviations obtained from two independent investigations. Statistics were calculated using unpaired student’s t test. PBMCs isolated from the patient and a healthy control were stimulated with TNF (200ng/ml) for 0, 5, 15, and 30 min. Levels of pIκBα and IκBα were investigated by western blotting (C). Protein sizes are marked on the left-hand side. Tubulin served as loading control
According to these findings, activation of NF-κB signaling in PBMCs upon TNF-stimulation showed reduced phosphorylation of IκBα (pIκBα) in the patient as compared to healthy controls (only one control shown) (Fig. 4C). Interestingly, pIκBα in response to TNF-stimulation was also reduced in PBMCs of her mother. Both, patient and mother, showed increased basal levels of IκBα. Of note, the patient was under treatment with TNF inhibitors at the time this investigation was performed.
Together these studies show an abrogated NEMO function with diminished response to TNF, but mostly retained TLR signaling in patient blood cells.
NEMO-Δex5 Results in Elevated IL-18 and CXCL10 ConcentrationsSince elevated cytokines have been observed in NEMO-NDAS patients, we measured the expression of 36 cytokines/chemokines in serum samples obtained from the patient, her mother and two controls. Patient samples were collected before start of treatment with anti-TNF agents. Expression of 13 cytokines could be detected on the membrane (Figure S3) represented by two spots per protein. Spot intensities were quantified by optical densitometry and normalized by an averaged background subtraction (Table S7). CCL5C5/C5aC, CD40L/TNFSF5, C5/C5a, CXCL1/GROα, CXCL10/IP-10, ICAM-1/CD54, IL-1ra/IL-1F3, IL-18/IL-1F4, MIF, Serpin E1/PAI-1 were significantly elevated in patient serum compared to mother and controls, of which CXCL10 and IL-18 were almost exclusively expressed in the patient (Fig. 5). An increased expression of the NF-κB-dependent chemokine CXCL10 has been reported before in NEMO-NDAS 21].
Fig. 5
NEMO-NDAS results in elevation of cytokines and chemokines. Levels of 36 cytokines were investigated in serum obtained from the patient, her mother, and control 1 and 2 using the Human Proteome Profiler Human Cytokine Array Kit (R&D Systems). Pixel densities were analyzed by optical densitometry and normalized on included background proteins. Shown are mean densities of 13 detectable cytokines. Statistics were analyzed using two-way Anova (* p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001)
Levels of total IL-18 were further investigated in patient serum samples collected during and between flares before start of treatment, as well as under treatment with adalimumab, and were compared to her mother and two controls. IL-18 level were elevated in patient samples at all time points (Table 1).
Table 1 IL-18 cytokine level in serum samplesExpression of NEMO-Δex5 is Caused by Random X Chromosome InactivationThe DNA methylation status of the X chromosomal IKBKG gene was studied in genomic DNA isolated from whole blood of the patient and both parents using nanopore sequencing (Fig. 6). In the mother, the allele carrying the IKBKG variant was methylated to 60% around the transcription start site, whereas no DNA methylation was detected within this region at the wildtype IKBKG allele. Thus, the results of the mother were in the technical range for assuming skewed X-inactivation. In contrast, both X alleles of the patient were methylated to approximately 50%/50% suggesting a random X-inactivation status at this locus (Fig. 6). Similar DNA methylation patterns could be observed in regions adjacent to IKBKG (Figure S4).
Fig. 6
The DNA methylation status of the IKBKG transcription start / promoter region is skewed to one X allele in the mother, but non-skewed in the patient (A) Exon 5 of IKBKG (chrX:154,560,348 − 154,560,622 in hg38) in patient, mother and father was analyzed by Oxford Nanopore Technologies (ONT) sequencing and results are displayed in the Integrative Genomics Viewer (IGV). The IKBKG variant c.613 C > T p.(Gln205*) is indicated by the red arrow and was found in the mother and the patient. (B) The DNA methylation pattern of patient, mother and father in the transcription start / promotor region of IKBKG (chrX:154,547,021–154,548,489 in hg38) was analyzed by ONT sequencing and is displayed in the IGV viewer. Wildtype (WT) allele and mutated (mut) allele of the patient (top panel) show similar DNA methylation patterns, whereas the WT allele from the mother is predominantly unmethylated (blue, middle panel). The mutated allele from the mother shows a DNA methylation of ~ 60%. The WT allele from the father (lower panel) is completely unmethylated (blue). No methylation = blue, methylation = red
A reversion mosaicism of the mutation in cellular subsets, as reported in several cases of NEMO deficiency [25], was excluded by Sanger sequencing (Figure S5).
Comments (0)