
Remember me


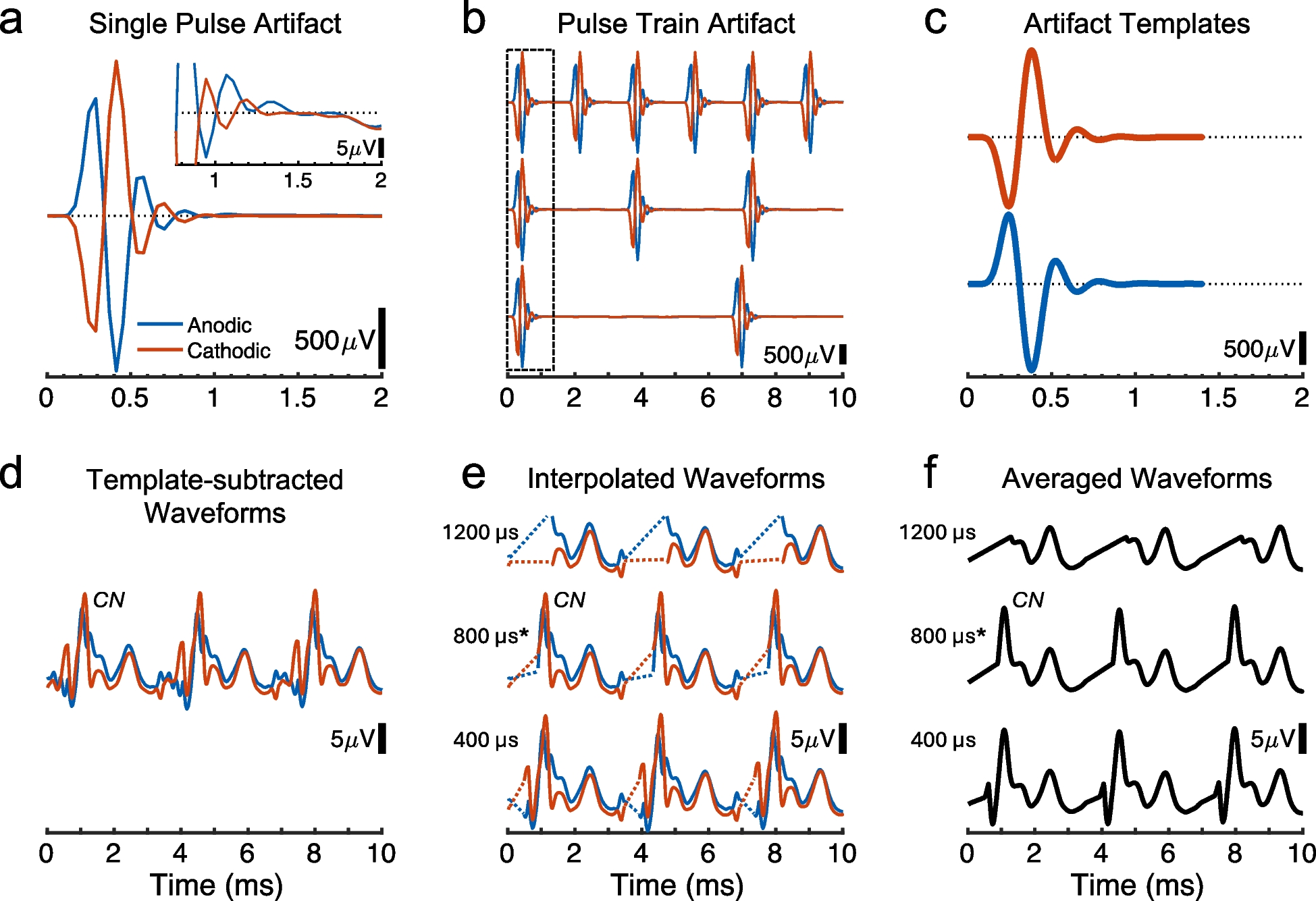
Measures of the eABR quantified the sensitivity of the auditory brainstem to single electrical pulses and the growth of response magnitude with increasing stimulus current. Measures of the eABR threshold at 2–3-week intervals assessed the stability of stimulation over the course of months after deafening and implantation. Figure 2a shows eABR waveforms from one cat recorded in response to single pulses from an IN electrode, 78 days post-implantation, in 2-dB steps of current; the background activity in a no-stimulus condition is also shown. The vertical gray bar indicates the 50–850-µs time range corresponding to blanking and linear interpretation for electrical artifact rejection (described in the “Methods” section). We attribute the first positive-going deflection, at peak latency ~ 1.2 ms, to the cochlear nucleus; the earlier compound action potential from the auditory nerve would have been lost in the artifact-rejection procedure. Waveform morphology varied among animals, recording laterality, and filter settings.
Fig. 2
eABR waveforms recorded from one cat, ipsilaterally to the stimulated ear, passband 300 to 1500 Hz. Stimuli were single biphasic pulses, 82 µs/phase. a The colored curves show the waveforms at current levels in 2-dB increments. no stim indicates background activity in the absence of overt stimulation. The vertical gray bar indicates the 50–850 µs time range corresponding to blanking and linear interpretation for electrical artifact rejection. b RMS voltages recorded during 1 to 4 ms after the pulse onset in 1- or 2-dB increments of stimulus current; current is given on the vertical axis, sharing the scale with Fig. 2a. Filled symbols denote recordings that were classified as above threshold according to the signal detection procedure described in the Methods. Cat AP, post-implantation day 78
Estimates of eABR thresholds were based on the root-mean-squared (RMS) voltage in the passband from 300 to 1500 Hz in the time window from 1 to 4 ms after pulse onset. Figure 2b plots the RMS values corresponding to the waveforms in Fig. 2a; stimulus current levels are given here on the vertical axis in 1-or-2-dB increments (aligned with the traces in Fig. 2a), and amplitude is given on a linear voltage scale on the horizontal axis. As detailed in the “Methods” section, we computed the cumulative d’ across successive current levels beginning with the no-stimulus condition. The filled symbols in Fig. 2b denote RMS values at which the cumulative d’ was ≥ 1, i.e., above threshold.
The growth of RMS of eABR amplitudes with increasing stimulus current is shown in Fig. 3a for each of the cats; data from test sessions ~ 70 days after implantation are shown for IN (blue) and CI (red) stimulation. Here, eABR amplitudes are given on the vertical axis as dB re the eABR amplitude at threshold for each cat, and current is given on the horizontal axis as dB re 1 µA. The symbol near the base of each curve (at 0 dB) indicates the eABR amplitude at threshold for that animal; the symbol near the top of each curve is explained in the following section. The maximum amplitudes of recorded eABRs were around 20–25 dB above the amplitude at threshold for both IN and CI stimulation. Note that the maximum amplitude that could be recorded for CI and, sometimes, for IN stimulation was practically limited by the myogenic (EMG) response that often was encountered at the highest stimulus levels. Despite the similar ranges of recorded amplitudes, the growth plots for IN and CI stimulation differed markedly in the stimulus ranges that elicited those response ranges. The thresholds for IN stimulation, expressed re 1 µA, are all lower than those for CI stimulation in the illustrated cases. Nevertheless, the amplitudes for both types of stimulation grow to maxima at around the same highest tested stimulus levels, around 50 dB re 1 µA. Consequently, the slopes of growth plots are shallower for IN than for CI stimulation. The difference in slopes is more evident in Fig. 3b, in which stimulus currents are expressed relative to the threshold for each plot. The thick lines plot the mean growth functions for IN (blue) and CI (red) stimulation. Slopes were shallower for IN than for CI stimulation.
Fig. 3
Growth of eABR amplitude (vertical axis) with increasing level (horizontal axis) of 82 µs/ph biphasic pulses. eABR amplitude is expressed as dB re the eABR amplitude at threshold. Each curve represents data from one animal studied ~ 70 days after implantation, blue for IN stimulation and red for CI stimulation. Symbols assigned to each cat (defined in the key) are maintained in this and in Figs. 4, 5, 6, 10, and 11. Symbols near the base of each curve indicate the eABR amplitude at threshold for each animal, and symbols near the top of each curve indicate the eABR amplitude at the stimulus level that was used for eFFR measurements. a eABR amplitudes shown at current levels expressed as dB re 1 µA. b Amplitudes shown at current levels expressed as dB re the threshold for each curve. The thick lines indicate the means across animals for IN and CI stimulation; the means shown by the thick lines were computed only across the range of suprathreshold levels that were tested in all the CI or all the IN animals
The distributions of eABR thresholds are shown in Fig. 4a. Means were 31.5 dB re 1 µA for IN stimulation and 43.0 dB for CI stimulation (two-sample two-tailed t-test: t = 7.2, p = 1.0 × 10−5, df = 12). Slopes are shown in Fig. 4b as dB amplitude/dB current. Mean slopes were 0.95 for IN and 2.0 for CI stimulation (t = 6.2, p = 4.9 × 10−5, df = 12). The relevance of growth-of-amplitude slopes to tonotopic spread of excitation is considered in the “Discussion” section.
Fig. 4
eABR level sensitivity for intraneural (IN) and cochlear-implant (CI) stimulation at ~ 70 days post implantation. a Thresholds. b Slopes of eABR growth quantified as eABR amplitude in dB/stimulus level in dB. In both panels, symbols are jittered in the horizontal dimension to aid in visualization. The assignment of symbols to cat codes is as in Fig. 3
The eABR thresholds for the various cats are shown as a function of days after implantation for IN (blue) and CI (red) stimulation (Fig. 5); filled markers indicate the animals that were implanted with CI + 1 devices. Two characteristics are most prominent. First, the distributions of IN and CI thresholds are largely non-overlapping, with CI thresholds higher than IN thresholds. The threshold difference was shown for single test sessions in Fig. 3a, but Fig. 5 shows that the difference persisted across months after implantation. Comparison of CI thresholds between CI-only and CI + 1 cases showed no consistent elevation of threshold in the CI + 1 cases, which alleviates a possible concern that the passage of IN electrodes through the fibers of the basal turn might have been injurious to those basal-turn fibers; the basal-turn fibers are the fibers activated by CI electrodes in the present study. Throughout this report, we assessed summary data on the test day for each animal closest to 70 days after implantation. That time point was chosen as a reasonably long period of chronic implantation that was achieved by all the cats in the study, even those that had hardware failures at later times.
Fig. 5
eABR thresholds versus days after implantation. Each symbol type indicates data from one animal keyed in the column at the right. Symbols are jittered by ± 1 day when necessary to avoid overlap. Colors and line styles indicate CI (red, dotted lines) and IN (blue, solid lines) stimulation. Filled markers indicate CI + 1 cats that were implanted with both CI and IN devices in the same ear in the same operation; CI and IN electrodes in those animals were tested in separate series of trials. The black arrow labeled BO (upper left) indicates the beginning of data for Cat BO for which the first three measurements were deemed invalid; those were the first test sessions in which such low thresholds were encountered, and the tested level range did not extend to sub-threshold levels. The vertical line at day 70 indicates the day closest to the test days that were used for two-sample two-tailed t-tests mentioned in the main text. The vertical line at day 183 indicates the target study longevity of 6 months
The second prominent characteristic of the data in Fig. 5 was that thresholds for individual animals showed little or no systematic variation across as long as 6 months (183 days) after implantation, even though those deafened animals received no stimulation during this time other than during the recording sessions, ~ 1–2 h every 2–3 weeks. Thresholds from the first three test days from cat BO were systematically overestimated and are illustrated but were excluded from across-day quantitative analysis. Among the seven tested IN electrodes, six reached our target longevity; we include Cats FO and BR among the 183-day count although connectors on those cats failed a few days short of the target, after days 155 (FO) or 177 (BR). Cat SC did not reach the 6-month mark because the skull-mounted connector became fouled, leading to marked increases in impedances after day 92. A two-way analysis of variance (ANOVA) for all the illustrated data (omitting the first three measurements from BO) showed a significant main effect of IN-vs-CI stimulation (F (1, 139) = 217, p < 10−15) but no significant effect of days after implantation in 14 blocks of 14 days (F (13, 139) = 1.1, p = 0.36). We take the main effect of stimulation type as indication that IN stimulation activated the auditory pathway at lower currents than did CI stimulation. The absence of significant effect of days after implantation indicates that there is no evidence for an effect of implantation on the functional health of the auditory nerves in these animals.
Electrically Evoked Frequency Following Response (eFFR)The eFFR provided a measure of transmission of TFS from the auditory-nerve stimulus to various brainstem, midbrain, and thalamocortical generators. We tested stimulus levels that were rather high in the dynamic ranges of eABR growth functions; the levels that were used for eFFR testing are marked by the symbols near the tops of the eABR growth plots in Fig. 3. The eFFR test levels were always set at least 2 dB below levels that elicited electromyogenic signals from facial nerve activation, when evident. Given the shallower slopes of IN eABR growth functions, that means that the highest practical stimulus levels were higher above threshold for IN than for CI stimulation. That is shown in Fig. 6a, which plots the distributions of levels that were tested, expressed as dB re threshold. Mean test levels re threshold were 19 dB for IN and 9.0 dB for CI stimulation (t = 6.4, p = 3.5 × 10−5, df = 12). Figure 6b plots the eABR amplitudes at the eFFR stimulus levels. Expressed in that way, the distributions of test levels for IN and CI overlap extensively, largely due to the shallower growth functions for IN stimulation. Mean amplitudes were 19 (IN) and 18 (CI) dB re threshold (t = 0.25, p = 0.80, df = 12). That result indicates that the stimulus levels that were chosen for eFFR testing were similar between IN and CI stimulation with respect to eABR amplitudes expressed relative to amplitudes at threshold.
Fig. 6
Stimulus levels used for eFFR measurements. a eFFR stimulus level re the single-pulse eABR threshold for each animal. b eABR amplitudes measured at the eFFR stimulus levels, given as dB re the amplitude of the eABR at threshold for each animal. Symbols are jittered in the horizontal dimension to aid in visualization. The assignment of symbols to cat codes is as in Fig. 3
Examples of time-folded eFFR waveforms from three series of responses to IN stimulation are given in Fig. 7. These waveforms were derived from samples at 50–500 ms re the onset of each pulse train; the response to the first 50 ms of each train was excluded to eliminate the first pulse, which tended to elicit a strong response irrespective of the rates of ensuing pulses. Each illustrated waveform is an average that is folded on a time range equal to the period of the pulse rate multiplied by the smallest integer multiple of that period that was ≥ 23.3 ms; 23.3 ms was the stimulus period of the lowest tested rate (43 pps). That means that the waveform for, say, the 152.6-pps pulse rate was folded on its period, 6.55 ms, times 4, which gave a total fold time of 26.2 ms. That folded waveform then was truncated to 23.3 ms, the length of the illustrated time axis. The colored waveforms represent pulse rates increasing from bottom to top; for clarity of presentation, the waveforms are shown only for rates < 400 pps. The waveform in black at the bottom of each column is the response to the 43-pps pulse rate, enlarged in amplitude to aid visibility of waveform features. The same features are present in all three cases but differ in prominence among recording-electrode placement and animals. The vertical gray bar in each panel shows the time range (from 50 to 850 µs) that was blanked and interpolated for artifact rejection; for clarity of presentation, the gray bar is shown only for the first pulse in each trace, although artifact rejection was applied to every pulse.
Fig. 7
Folded eFFR waveforms, passband 50–1000 Hz. Waveforms recorded from two IN cats were elicited from 50 to 500 ms after the onsets of electric pulse trains, with the pulse rates indicated on the vertical axes. The time base for each pulse rate is folded on the period of that rate times the smallest integer multiple that totaled ≥ 23 ms. The vertical gray bar in each panel shows the time range (from 50 to 850 µs) that was blanked and interpolated for artifact rejection; for clarity of presentation, the gray bar is shown only for the first pulse in each trace, although artifact rejection was applied to every pulse. The bottom trace in each panel, in black, shows the response to the lowest pulse rate (43 pps) expanded in the amplitude dimension. a Cat AP (78 days after implantation); ipsilateral recording. b Cat AP (day 78); contralateral recording. c Cat SC (day 79); contralateral recording. CN, cochlear nucleus; SOC, superior olivary complex; IC, inferior colliculus; MGB, medial geniculate body; TC, thalamocortical projection; CTX, primary auditory cortex
The waveforms in Fig. 7a were ipsilateral recordings from cat AP. The enlarged (black) trace for the 43-pps rate shows a prominent vertex-positive deflection at ~ 1.30 ms, which, based on the latency, we attribute to the cochlear nucleus (CN); the compound action potential from the auditory nerve at latency < 1 ms was lost in the artifact-reduction procedure. The CN deflection was followed by another positive-going deflection, at ~ 2.7 ms, and a negative-going deflection at ~ 3.5 ms, which we attribute to the superior olivary complex and possibly additional brainstem generators (denoted here collectively as SOC). The CN deflection can be seen at all the illustrated pulse rates (shown in colors). At the higher rates, the CN deflection increases slightly in latency, from 1.33 to 1.48 ms, and broadens, both of which we attribute to habituation at the high pulse rates. There is a positive deflection at latency ~ 5 ms, which is within the range of latencies of electrically evoked spike activity in the ICC [10, 21]. The waveforms in Fig. 7b are the contralateral recordings from the same cat (AP) as in Fig. 7a, c shows contralateral waveforms from cat SC. Viewed from the contralateral side, the CN deflection is negative-going and smaller than in the ipsilateral recording, but the IC deflection at ~ 5 ms is especially prominent, particularly in Fig. 7b. The IC deflection in this laterality is evident in the colored waveforms at pulse rates up to ~ 200 pps or higher. There is a broad positive deflection centered at ~ 9–10 ms at the lowest pulse rate, more prominent in Fig. 7c. That latency is consistent with activity in the medial geniculate body (MGB; [22]).
Figure 7b, c each show two deflections that likely originated in the auditory cortex. The first is a negative deflection (labeled TC) centered at 12 ms (in Fig. 7b) or 15 ms (Fig. 7c), which likely reflects activity in thalamic input layers of primary auditory cortex [23]. That first deflection likely also includes the thalamocortical excitatory post-synaptic potentials. The second deflection (labeled CTX) is positive at 19 ms in Fig. 4b and > 23 ms in Fig. 4c, similar to group delays of spike activity in the unanesthetized primary auditory cortex of cats in response to acoustic click trains ([24] and unpublished data from that study). The sizable delay between the thalamic input and group delays of spike activity agrees with the intra-cortical temporal filter delay of 7.5 to ~ 14 ms between first-spike latencies and group delays described by Eggermont for cat acoustic click trains [25]; similar delays between cortical first-spike latencies and group delays were reported for CI stimulation in guinea pigs [26]. The colored lines in Fig. 7c show the MGB deflection synchronizing to pulse rates > 108 pps, whereas the TC deflection is evident only to about 55 pps.
At the low pulse rate shown in black in Fig. 7, one can easily identify waveform deflections arising from putative generators. At higher pulse rates, however, long-latency deflections are overtaken by short-latency responses to following pulses. This is particularly evident for the CTX deflection in Fig. 7b, and the MGB and TC deflections in Fig. 7c. The overlap between long-latency responses to earlier pulses and short-latency responses to later pulses means that the eFFR recorded at rates higher than ~ 50 pps is given by the sums of multiple brainstem, midbrain, and thalamocortical generators.
The sum-of-generator concept has been explored in normal hearing (cat: [18]; rabbit: [19]; human: [20]) and CI hearing (cat: [11]). Here, the sum-of-generators characteristic of eFFR recordings was evident in the transfer functions from stimulus pulse rates to eFFR recordings. Amplitude transfer functions from IN (Fig. 8a) and CI (Fig. 8c) stimulation cases exhibited prominent peaks and dips, which are characteristic of a sinusoid added to itself with multiple delays. Within particular CI or IN electrodes in individual cats, the magnitudes of peaks and dips could vary considerably across test sessions. Nevertheless, the pulse rates corresponding to peaks and dips in amplitude transfer functions were remarkably consistent across various test sessions within each cat. For example, we identified the pulse rates at three dips in the amplitude transfer function in the ~ 70-day test session for each cat and compared those rates with corresponding rates in the ~ 100-day test session. We scored a “hit” whenever the rates at 70 and 100 days differed by no more than 32 pps, which was the smallest pulse-rate increment that was tested across all ranges of pulse rates. Hits were observed in 39 (92.9%) of 42 cases (i.e., three dips in each of seven CI cases and seven IN cases = 42); the median magnitude of error across hits and misses was 0 pps (interquartile range 0 to 15.5 pps). The few misses were instances in which dips were very shallow in one or both of the test sessions. The variation in transfer functions was considerably greater across the various cats. We evaluated the inter-cat differences by testing the same 42 conditions (three dips × 14 cases) with 200 random permutations of the day-70 day sessions from one cat and the day-100 sessions from a different cat. That yielded substantially lower hit rates: median = 61.9%, 95th percentile = 71.4%.
Fig. 8
Pulse-rate transfer functions: amplitudes (a, c) and cumulative phases (b, d) are the Fourier amplitudes and phases at the component closest to each pulse rate. The vertical dashed lines passing from a to b and from c to d indicate nulls in amplitude at which phase was ambiguous or the rate of growth of phase lag declined. The cumulative phases each are fit with three or four line segments using piecewise linear regression. Group delays, indicated in ms, were computed from the slopes of the line segments. The right-most end of each line segment indicates the pulse-rate cutoff, which was the maximum pulse rate at which the eFFR was dominated by a generator having the corresponding group delay. In these examples, the pulse-rate cutoff for a putative midbrain generator having a latency of ≥ 4 ms is indicated by the dotted vertical lines at 370 pps (a) and 200 pps (b). a, b: Cat BO day 57; c, d: Cat XE day 73
The phase transfer function from the same cases as Fig. 8a, c are shown in Fig. 8, d. Vertical dashed lines between upper and lower panels show instances in which phase values were poorly defined at pulse rates corresponding to nulls in the amplitude transfer function. Phase transfer functions typically showed multiple segments of relatively straight increasing phase lag with increasing pulse rate. In the example in Fig. 8b, from an IN case, piecewise linear regression could fit most of the phase values with three line segments, running from pulse rates of 48 to 108 pps, 122 to 244 pps, and 271 to 370. The slope of each of those segments divided by \(2\pi\) yielded the corresponding group delay, equal to 12.8, 6.9, and 4.5 ms in this example. We assume that the group delays reflect the neural latencies of particular generators in the auditory pathway. The group delays here are consistent with scalp recordings dominated by thalamocortical (12.8 ms), midbrain to thalamic (6.9 ms), and IC (4.5 ms) generators. In this example, 370 pps, marked with a blue dotted line, was taken as the cutoff rate for synchrony dominated by midbrain levels of the auditory pathway. In the example from a CI case (Fig. 8b, d), the phase transfer function could be fit well with four line segments. The segment having a group delay consistent with a midbrain generator, 4.8 ms, only reached to 200 pps (red dotted line).
Based on the piecewise linear regression analysis of phase transfer functions, we compiled across animals the group delays of the generators that dominated the response to each pulse rate. The distributions of those group delay values are shown in Fig. 9 for seven IN cases (in blue) and seven CI cases (in red). Interquartile ranges (vertical bars) and medians (short horizontal bars) are given for each pulse rate and IN and CI device. We limited this analysis to pulse rates < 500 pps because the eFFR at higher rates was dominated by short-latency brainstem activity in which group delays were poorly defined in the piecewise regression analysis. The horizontal rows of numbers in the figure gives the p values for two-sample t-tests comparing the group delays for CI-versus-IN stimulation at each pulse rate; p values at rates < 150 pps were all > 0.14 but are omitted for clarity. The t-tests showed significant differences at rates of 321 and 370 pps, but none of the p values was significant after Bonferroni correction for 19 repeated observations.
Fig. 9
Distributions of group delays as a function of stimulus pulse rate. Data are from the test days closest to 70 days after implantation compiled across cats. The analysis included seven IN cases and seven CI cases, but group delays were not measurable at all pulse rates for all cats. For that reason, there are four to seven (median = 6) cats included for each IN condition and two to seven (median = 6) for the CI conditions. Each pair of red (for CI stimulation) and blue (for IN) vertical lines represents the interquartile range of group delays computed for each pulse rate, and the short horizontal lines indicated the median values. The values printed across the top are p values for two-sample t-tests at each pulse rate; there was no assumption that the CI and IN distributions had equal variance. The p values at rates < 150 pps are omitted for clarity, but all were > 0.14
A two-way ANOVA of the data in Fig. 9 across the 19 pulse rates from 68 to 421 pps showed a main effect of pulse rate (F (18, 174) = 24, p < 10−15) but no significant effect of CI-versus-IN stimulation (F (1, 174) = 0.43, p = 0.51). The main effect of longer group delays at lower pulse rates is expected because midbrain and forebrain auditory structures having long latencies (and long group delays) are known to synchronize only to relatively low rates [10, 24, 27, 28]. The absence of a significant main effect of CI-versus-IN stimulation seemingly is contrary to our expectation of enhanced brainstem synchrony to IN stimulation, which is based on invasive ICC unit recordings [10]. The present analysis, however, includes a range of high pulse rates in which eFFR amplitudes were low and for which the response is dominated by peripheral sites for which the group delays would not be expected to differ between stimulus type. Also, there was a range of low rates, ≤ 150 pps, for which synchrony of thalamocortical generators would be expected to show a ceiling for either electrode type. The figure shows, however, that group delays consistently are higher for IN than for CI stimulation across pulse rates around 300 pps and group delays > 4 ms, which are relevant to synchronized unit activity in the ICC [10]. Nearly all the units recorded in the previous ICC study showed first-spike latencies and group delays from 4 to 9 ms, and the difference in ICC synchrony between CI and IN stimulation became conspicuous at pulse rates around 300 pps and higher. For those reasons, we define eFFR group delays ≥ 4 ms as a region of interest in the present study and henceforth will use the midbrain sync cutoff to refer to the highest stimulus pulse rate at which the group delay was ≥ 4 ms. In Fig. 9, the eight pulse rates in which the interquartile range for IN and/or CI stimulation included 4 ms ranged from 218 to 394 pps. If we limit our two-way ANOVA to that range of rates, we observe significant main effects of pulse rates (F (7, 76) = 4.6, p = 0.00024) and stimulation type (F (1, 76) = 23, p = 7.8 × 10−6). The main effect of stimulation type across that limited range of pulse rates accords with the previous result that substantially more ICC neurons synchronized at pulse rates of ≥ 300 to IN compared to CI stimulation [10].
The data in Fig. 9 were collected from the single test session for each cat that was nearest to 70 days after implantation. For the goals of the study, it was of interest to monitor possible long-term changes in midbrain sync cutoffs. Figure 10 shows midbrain sync cutoffs as a function of days after implantation for IN (blue) and CI (red) stimulation. With few exceptions, eFFR at group delays consistent with midbrain generators (i.e., ≥ 4 ms) was present at higher pulse rates for IN than for CI stimulation. The trends across days made a few large swings, which we attribute to difficulty in interpreting phase spectra around dips in amplitude spectra and, in some cases, difficulty in placing electrodes to record a dominant midbrain generator. Nevertheless, there were no consistent long-term trends across post-implantation time. A two-way ANOVA of eFFR cutoff showed a significant main effect of IN-versus-CI stimulation type (F (1, 115) = 114, p < 10−15; means across all days, 367 pps for IN and 241 pps for CI), whereas there was no significant main effect of days after implantation compiled in 14 groups of 14 days (F (13, 115) = 0.87, p = 0.59); similar significance of device type (F (1, 106) = 94, p < 10−15) and lack of significance of days after implantation (F (13, 106) = 0.84, p = 0.62) were observed when the analysis was repeated after excluding the test days that were used in Fig. 9. We take the main effect of device type, that midbrain synchrony cutoffs were higher for IN than for CI stimulation, as indication that IN stimulation activated brainstem pathways that transmit TFS more effectively than do pathways activated by CI stimulation. As in the case of stable eABR thresholds, the lack of significant effect of days after implantation fails to support any hypothesis of an ill effect of implantation and stimulation on the maintenance of auditory-nerve function.
Fig. 10
Midbrain sync cutoffs as a function of days after implantation. The midbrain sync cutoff is the highest pulse rate at which the analysis of the eFFR phase transfer functions showed a group delay ≥ 4 ms. Markers are jittered by ± 1 day when necessary to avoid overlap. Filled markers denote CI + 1 implants. Blue color and solid lines denote IN stimulation; red color and dotted lines denote CI stimulation. Other details as in Fig. 5
Temporal Acuity Maintained After DeafeningIn a previous study, we found that cats that were deafened and left unstimulated for ~ 6 months showed a marked degradation of temporal acuity, as measured by loss of phase-locking of single- and multi-unit activity in the ICC [13]. Animals in the present study were similarly deaf for ~ 6 months but were stimulated unilaterally through an implanted electrode for ~ 1–2 h at 2–3-week intervals. Those animals showed no systematic loss of temporal acuity as measured by the eFFR. In a subset of those animals, we tested temporal acuity in terminal experiments more than 80 days after cessation of routine stimulation. Those animals were sedated, and CI and IN electrodes were implanted acutely, one at a time, in the formerly un-implanted ear. Figure 11 shows the eFFR midbrain synchrony cutoffs from those seven cats. The red or blue symbol indicates the value that was recorded with the chronic stimulating electrode in the last routine recording session; each of these animals had only one or the other type of chronic electrode. The red and blue horizontal lines indicate values obtained in the terminal recording session with the acutely implanted electrodes. In every case, the acute IN stimulation yielded midbrain synchrony at higher rates than did acute CI stimulation (pairwise t (6) = 6.8, p = 0.00051). This replicates the effect of device on midbrain synchrony, observed in the analysis of the data presented in Figs. 9 and 10, with a fresh data set. The maximum synchrony rates from the acute CI and IN stimuli after > 80 days of no stimulation generally were lower compared to the chronic stimulation in those animals (pairwise t (6) = 3.4, p = 0.014). In the two exceptions, at 81 and 126 days unstimulated, the acute CI value was lower than the chronic CI value, although the acute IN value was higher than both CI values. The results of these terminal experiments support the view that temporal acuity was sustained in deafened animals by routine electrical stimulation and that acuity was degraded after a period of > 80 days of no stimulation.
Fig. 11
Loss of temporal acuity after > 80 days of no stimulation. Each vertical triad of symbols represents data from one animal, with the horizontal dimension plotting the stated number of days after the cessation of routine electrical stimulation. The symbols, corresponding to the cat codes defined in Fig. 3, indicate the midbrain synchrony cutoff obtained with the chronic electrode in the last routine recording session, either IN (blue) or CI (red), and the horizontal lines indicate the values obtained in a terminal experiment with acute implantation of IN (blue) and CI (red) electrodes in the formerly un-implanted ear
Comments (0)