
Remember me



Traumatic brain injury (TBI), the dominant type of head injury, subjects more than 10 000 000 people to death or hospitalization every year globally. TBI is mainly caused by falls, traffic incidents, and beats [1]. Notably, TBI has emerged as a dominant cause of morbidity and mortality. The significant expenses on clinical caring of TBI patients and related socioeconomic issues will severely stress the healthcare system and society [2]. Much research in the past ten years suggests that mesenchymal stem cells (MSCs) are potential and efficient for brain injury treatment, in both testing models of TBI and potentially in clinic [3–6]. Despite such findings, the underlying roles of MSCs in post-TBI brain tissue repair and function recovery may not relate to the cell replacing impacts, but to the secretion-based paracrine impacts [7–9].
Exosomes are lipid bilayer membrane vesicles in diameter of ≤ 150 nm and are produced by diverse cells, such as MSCs and cancer cells [10,11]. Exosomes with abundant endosome-originating parts (e.g. microRNAs, mRNAs, proteins) are often investigated by marker proteins, transmission electron microscopy (TEM), and nanoparticle size examination [12]. Compared with the parent cells, MSC-derived exosomes (MSCs-Exo) have less immunogenicity due to limited or no proliferating ability and can be safely kept and moved without function loss [13]. The effectiveness of MSCs-Exo was verified in animals with TBI [5,14]. The function mechanism of MSCs-Exo is yet unknown but can be attributed to the post-injury inhibition of inflammatory reactions. Microglia, a significant factor in aggravating and modulating neuroinflammation, can impose a double-edge impact on TBI neurogenesis [15]. In addition to inflammatory molecules [interleukin (IL)-1β/6, tumor necrosis factor (TNF)-α] that are detrimental to neurogenesis, microglia can also secrete anti-inflammatory factors (e.g. transforming growth factor β, IL-4/10) that are beneficial to neurogenesis [16,17]. Reportedly, the use of MSCs-Exo led to neuroinflammation reduction and remarkable functional restoration in a rat TBI model [18]. Exosomes produced from MSCs may be able to limit neuroinflammation and encourage brain repair by controlling the morphologies of nearby microglia, according to some researchers [19]. Because of their safety and minimal immunogenicity, while yet displaying the same immunomodulatory and regenerative abilities as their mother cells, MSC-derived exosomes have a great deal of promise for cell-free therapy [20]. Exosomes have been utilized therapeutically to decrease neuroinflammation following TBI. It has been demonstrated that MSC-derived extracellular vesicles suppress microglia activation [21]. Thus, exosomes can relieve TBI by changing microglial roles in vivo, which is not yet clarified, however.
Nucleotide-binding oligomerization domain-like receptors (NLRs), one subgroup of pattern recognition receptors, are critically involved in inherent immune/ inflammatory reactions by producing inflammasomes [22] that are distributed discrepantly in the brain. The NLR family pyrin-domain-containing 3 (NLRP3) is mainly located in microglia [23]. NLRP3 inflammasome, a cytosolic signaling compound, is composed of NLRP3, the apoptosis-related speck-shaped protein with a caspase recruiting zone, and pro-caspase-1 [24]. NLRP3 inflammasome was up-expressed in animal models and TBI patients [23,25], and its isolation or knockdown weakened inflammatory reactions and enhance neurological roles in TBI rodents [26,27]. Studies on the control of NLRP3 by exosomes and inflammatory processes have exploded recently [28–30]. For instance, Yan et al. [31].discovered that exosomes produced by umbilical cord MSCs (UMSC-Exo) reduced the amount of cleaved caspase-1 produced, which in turn reduced the release of IL-1 and IL-18 and pyroptosis. circHIPK3 produced by UMSC-Exo down-regulated miR-421, leading in enhanced expression of fork head box class O 3a, which might suppress NLRP3 activation. Exosomes can influence immunity in disease states by keeping an eye on gp130/STAT3 signaling, the TLR4/NF-B/NLRP3 inflammasome, and programmed cell death-1 [32]. Herein, we discussed the relationship between the exosomes and NLRP3. Therefore, the NLRP3 inflammasome is a new potential target for exosomal treatment of TBI.
Adipose-derived stem cells (ADSCs) offer a microenvironment for cell survival by generating neurotrophic, neuroprotective and anti-inflammatory factors, and reduce the formation of stress-related proteins, reactive oxygen, and pro-inflammatory factors [33–35]. Hence, it is suggested that ADSCs-derived exosomes (ADSCs-Exo) may also reduce neuroinflammation and facilitate functional recovery from TBI. Herein, the composition of ADSCs-Exo and the impact on microglial activation were explored in an in-vitro TBI model. Also, the potential mechanism was discussed with an in vivo mouse TBI model.
Materials and methods Ethics and approvalsA total of 62 rats were used in the study. This is due to the fact that post-trauma motor and cognitive function did not significantly differ between female and male SD. In contrast, the immunological response was more stable in female SD. This experiment involved using female SD rats [36–38]. All animal experiments were approved by the Animal Care and Use Committee of East China Normal University.
ADSC separation and characterizationADSCs were obtained as reported [39]. Female Sprague–Dawley (SD) rats (6–8 weeks old, 180-220 g) were used for our experiment. The inguinal adipose tissue was cleaned with a phosphate buffer solution (PBS). After 5 min of centrifugation at 1200 rpm, the supernatant was isolated from the cell suspension. The pellets were cautiously suspended with PBS (10 mL) and centrifuged again. The pellets without the supernatant were dissolved again in an erythrocyte lysis solution (10 mL) and cultured for 10 min at ambient temperature. After red blood cells were lysed, the cell solution was passed via a 100-lm nylon filter to clean off cell debris. The cells were gathered and sown on culture plates with Dulbecco’s modified Eagle medium (DMEM), 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 mg/mL streptomycin. After 24 h, non-adhering cells were discarded, a new full medium was added and changed every third day then. Multipotency of separated cells was clarified by adipogenic differentiation, which was conducted with the cells cultured for 2 weeks in DMEM supplemented with 10% FBS, 0.5 mM isobutylmethylxanthine, 1 μM dexamethasone, 10 μM insulin and 200 μM indomethacin media and dyed by Oil Red O. DMEM medium (SH30081.01) and FBS (SH30088.02) were from Hyclone. Penicillin (5161), streptomycin (85886), isobutylmethylxanthine (I5879), dexamethasone (D1756), insulin (I1507) and indomethacin (I7378) were from Sigma.
ADSC-Exo isolation and characterizationCells were planted with serum at 37 °C and 5% CO2 until reaching a subconfluent status (80–90%). Then the cells were slowly cleaned with PBS three times and cultivated for 48 h in serum-free DMEM. The cell-conditioned solution was collected and centrifuged successively at 300 × g, 2000 × g (both 10 min), and 10,000 × g (30 min) to discard the cell residues and debris. The exosomes were pelleted under an SW32 Ti shaking bucket rotor from the supernatant and ultracentrifuged at 100,000 × g for 70 min. To eliminate protein contamination, the pellets were suspended again in PBS, pooled, and ultracentrifuged again under the same condition. The final exosome pellets were suspended again in PBS until used.
Diameter distribution of ADSCs-Exo was analyzed under ZetaView (Particle Metrix GmbH, Meerbusch, Germany). Morphology was recorded by a Hitachi H7500 TEM device (Tokyo, Japan). Surface markers CD63 (ab193349, Abcam) and CD81 (ab109201, Abcam) encapsulated into exosomes were examined by Western blot.
Stimulation with lipopolysaccharide and co-culture with ADSCs-Exo for primary microgliaPrimary microglia were separated and purified from rat cortices as described before [40]. The microglia were acquired from the astrocytic single layer by beating the flasks 10 to 20 times and planted to poly-d-lysine-coated culture plates until used. In 150-cm3 culture flasks, 5 107 cells were seeded with DMEM containing 10% heat-inactivated FBS, 2 mM l-glutamine, 1 mM sodium pyruvate, 100 mM non-essential amino acids, 50 U/ml penicillin, and 50 g/ml streptomycin. Microglial cells were exposed to 100 ng/ml lipopolysaccharide (LPS; SigmaAldrich) and cultured with 5 or 10 μg/ml ADSCs-Exo for 24 h, so as to evaluate the effect of ADSCs-Exo on LPS-stimulated primary microglial cells. Cells processed without either LPS or exosomes were set as controls. LPS (L3012), l-glutamine (G2150), sodium pyruvate (P2256), and non-essential amino acids (TMS-001) were from Sigma.
ELISA of pro-inflammatory cytokinesDifferent treatments were used during the 24-hour cultivation of primary microglia cells. The culture supernatant was then taken out and spun at 3000 rpm for 20 min. Finally, using the appropriate ELISA kit per the manufacturer’s instructions, the cellular abundances of TNF-α (ab236712, Abcam), IL-1β (ab255730, Abcam), and IL-6 (ab234570, Abcam) were found in the medium.
Construction of TBI rat model and application of exosomesUnder anesthesia via intraperitoneal administration of 40 mg/kg pentobarbital sodium (1%), the skull of each animal was exposed via sagittal cutting and fascia reflection at the scalp midline. Then a 4.0-mm-diameter craniotomy on the right hemisphere was made between the lambda and bregma and 3.0 mm from the sagittal suture. The skull flap was also discarded. A 2.5-mm-diameter pillar was positioned at the hit site to release a 40-g weight dropping from 10 cm high along a stainless-steel string, and then was stricken. After the injury, the scalp cut was sewn. The sham rats were received craniotomy only, except for contusion.
Rats were randomly classified into 3 groups: sham, TBI, or TBI+ADSCs-Exo (n = 7/group). ADSCs-Exo (200 µg precipitated in 200 µL of PBS) was intravenously applied over 5 min via the tail vein at 24 h after the preparation of TBI. Equal volumes of PBS (200 µL) were administered in the TBI and sham groups.
Evaluation of neurological outcomeThe modified neurological severity score (mNSS) integrates sensory (vision, tactile, and proprioceptive), reflex, and motor (muscle status, abnormal motion) assays and is extensively adopted before. The mNSSs of each rat were detected at 0, 1, 4, 7, 14, 21, 28 and 35 days after TBI [41]. Neurological function was scored from 0 (normal) to 18 (maximum deficit). Each abnormal conduct or lack of a tested reflex was assigned one point. Hence, a larger score stands for a severer injury.
Foot faults were tested [42] at the same time points as mNSS. The rats can walk on a grid. Foot fault was defined as the fall or slip of a paw between the wires at each weight-bearing step. Totally 50 steps for the right forelimb were observed.
ImmunofluorescenceCells were fixated in 4% paraformaldehyde, and cleaned two times in 1× PBS. The primary antibody was anti-Iba1 (ab178846, 1 : 100, Abcam). After nuclei were dyed with DAPI (ab104139, 1 : 1000, Abcam), the cells were examined by fluorescence microscopy. All images were processed on ImageJ (software version 1.52; National Institutes of Health, Bethesda, MD, USA) [43].
Western blotThe total protein of the cortical tissue close to or in the vicinity of the damage site was taken for assay. Brain tissues were treated with a RIPA protein lysis solution and total proteins were harvested via centrifugation. Western blot was carried out as per standard steps. Primary antibodies were NLRP3 (ab263899, 1 : 1000, Abcam), Caspase-1(ab286125, 1 : 1000, Abcam), and β-actin (ab8227, 1 : 5000, Abcam). Membranes were observed by an enhanced chemiluminescence Western blot device (Thermo Fisher Scientific). Gray values were tested on Image J (Rawak Software, Germany).
Statistical analysisGraphPad Prism Software 8.0 (GraphPad Software, USA) was used to conduct the statistical analysis. All data were analyzed using Student’s t-test for two groups, one-way analysis of variance (ANOVA), or two-way ANOVA with Bonferroni’s multiple comparison test for more than two groups. The data are all reported as the mean ± SD. When P < 0.05, differences between means were regarded as statistically significant.
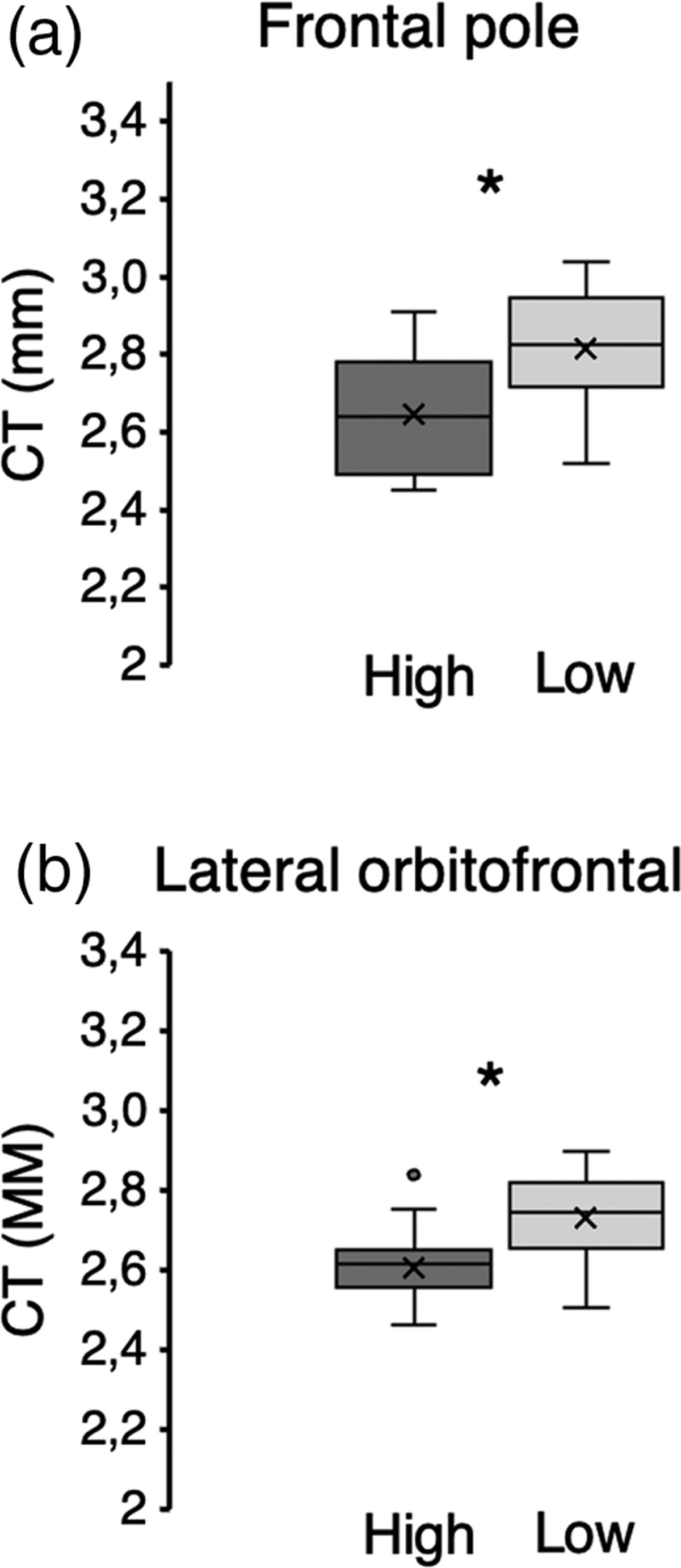
Results Characterization of ADSCs-ExoThe ADSCs are plastic-adhering and look like fibroblasts (Fig. 1a). The cells were processed by adipogenic solutions and confirmed by Oil Red O dyeing of lipid drops (Fig. 1b). The ADSCs-Exo were comprehensively characterized by TEM and Western blot. ADSCs-Exo exhibits a round shape under TEM (Fig. 1c), and CD63 and CD81 are positive in ADSCs-Exo as per Western blot (Fig. 1d), which both confirms the existence of exosomes. The above results reveal the triumphant isolation and recognition of ADSCs-Exo.
 Fig. 1:
Fig. 1: Characterization of ADSCs and ADSCs-Exo. (a) ADSCs as monolayer fibroblast-like cells. (b) Lipid vacuoles stained with Oil red O after adipocytic division. (c) TEM of ADSCs-Exo. (d) Western blotting was used to identify exosome-specific markers (CD63 and CD81). Results are representative of n = 3 independent experiments.
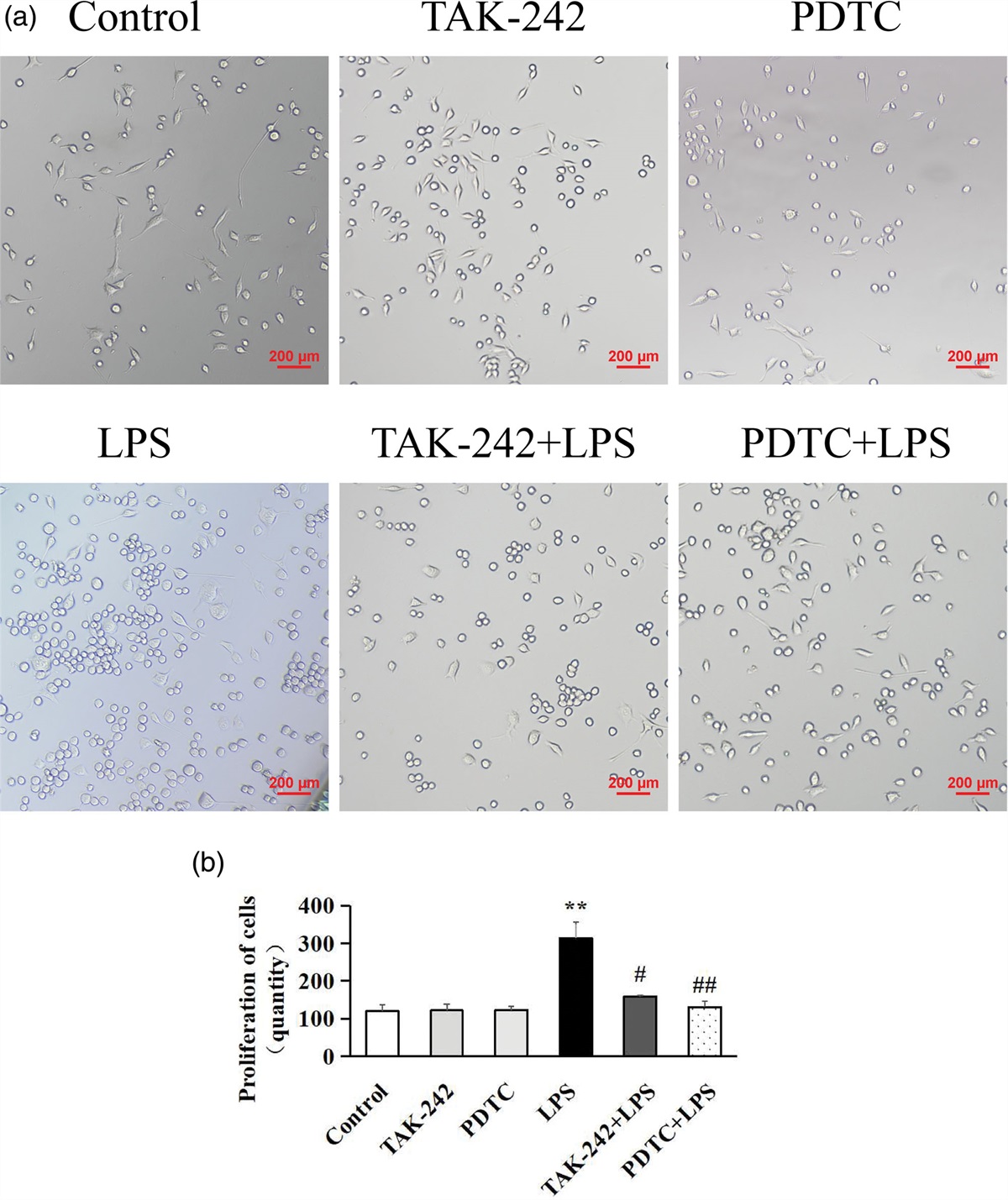
Exosome internalization reduces LPS-initiated generation of pro-inflammatory molecules in microglial cellsThe direct impact of ADSCs-Exo on pro-inflammatory molecule alteration was further verified in the primary microglia (Fig. 2a–i). The LPS-induced morphology of microglial cells is altered by ADSCs-Exo. LPS-treated (100 ng/ml) microglial cells showed long thin processes extending from the cell body, in contrast to the control group. The number of long thin processes extending from the cell body in cells cultured with ADSCs-Exo (10 µg/ml) followed by LPS treatment was decreased, and these cells had the similar appearance as the control group.
 Fig. 2:
Fig. 2: Functional influence of ADSCs-Exo on LPS-induced microglia in vitro. (a–i) Primary microglia were co-cultured 24 h in three groups: PBS control, LPS (100 ng/ml LPS), and ADSCs-Exo (100 ng/ml LPS + 10 µg/ml ADSCs-Exo). Primary microglia detected through specific markers Iba1 (green) by immunostaining analysis. Levels of (j) TNF-α, (k) IL-6 and (l) IL-1β in the cell supernatant of each group. The pro-inflammatory molecules were detected by ELISA. The data are expressed as the mean ± SD (n = 7/group). *P < 0.001 vs. the control group; #P < 0.05, ##P < 0.001 vs. the LPS group.
In the microglia, 100 ng/ml LPS stimulation initiated the secretion of IL-1β/6 and TNFα. Compared with LPS-processed cells (Fig. 2j), ADSCs-Exo restricted the LPS-caused TNFα production by 5 µg/ml (1445 pg/ml) and 10 µg/ml (913 pg/ml) (both P = 0.001). Additionally, ADSCs-Exo decreased the LPS-induced IL-6 secretion by 5 µg/ml (290 pg/ml; P = 0.035) and 10 µg/ml (178 pg/ml; P = 0.001) (Fig. 2k). Finally, ADSCs-Exo reduced the LPS-caused IL-1β formation by 5 µg/ml (503 pg/ml) and 10 µg/ml (485 pg/ml) (both P = 0.001) (Fig. 2l). Such results indicate that ADSCs-Exo inhibit pro-inflammatory responses in primary microglia by LPS.
ADSCs-Exo application significantly enhances post-TBI sensorimotor functional restoration in ratsThe mNSSs on Day 1 post-TBI were near 12 in all rats, implying no obvious difference in the neuro-deficits among the TBI rats at baseline. When compared to day 1 post-injury, the mNSS score in the ADSCs-Exo group animals decreased significantly over time, starting at Day 7 and continuing through Day 35 (P < 0.05), indicating a considerable improvement in mNSS following TBI (Fig. 3a). Before the assignment of treatments (PBS or ADSCs-Exo), there was no discernible difference in the footfault tests between the TBI groups on day 1 post-injury. In the ADSCs-Exo group animals, a substantial decrease in the number of foot faults was seen over time, commencing on day 14 and continuing until day 35 compared to day 1 post-injury (P < 0.05), indicating the presence of a considerable recovery in foot faults following TBI (Fig. 3b). Thus, ADSCs-Exo administration contributed to sensorimotor functional recovery in the TBI rats.
 Fig. 3:
Fig. 3: Administration with ADSCs-Exo significantly improved sensorimotor functional restoration as detected by modified neurological severity score (a), right forelimb foot fault (b) in rats after TBI. *P < 0.05 vs. PBS group. Data were expressed as mean ± SD (n = 7/group).
ADSCs-Exo inhibit NLRP3-mediated inflammasome activity after TBIPost-TBI stimulation of NLRP3 inflammasome and the inhibitive effect of ADSCs-Exo on NLRP3 were examined by detecting the expressions of the parts of NLRP3 inflammasome and cleaved activated caspase-1 using Western blot (Fig. 4a). NLRP3, caspase-1 were decreased in ADSCs-Exo rats after injury compared to the TBI group, as shown in Fig. 4b and c (P < 0.05). This implies that NLRP3 activation and its downstream activities are blocked by ADSCs-Exo downregulation.
 Fig. 4:
Fig. 4: Effect of ADSCs-Exo on NLRP3 inflammasome expression post-TBI. (a) Western blot of NLRP3 and caspase-1 at 24 h post-TBI, and relative expressions of (b) NLRP3 and (c) caspase-1 in pericontusional cerebral cortex. Data were expressed as mean ± SD (n = 7/group). *P < 0.001 vs. sham group; #P < 0.05 vs. TBI group.
DiscussionMSCs-Exo can be applied in ideal cell-free treatment of TBI. Investigation into the effects and mechanisms of ADSCs-Exo on TBI reveals that ADSCs-Exo can resist inflammation in TBI. The exosomes can lower the formation of pro-inflammatory factors by microglia in vitro following LPS activation. Vein administration of ADSCs-Exo in vivo is an efficient cure for neurological function recovery by reducing NLRP3 inflammasome activation.
Testing models and clinical applications of TBI indicate that MSCs are a promising and prospective treatment for brain injuries. However, as reported recently, the main mechanisms of MSCs involved in post-TBI cerebral reconstruction and functional repair may be the paracrine impacts of secretion-related factors (e.g. MSCs-Exo) rather than cell replacing effects. Exosomes comprise of proteins, nucleic acids, miRNAs, lipids, and mRNAs [44]. MSCs-Exo can relieve neuroinflammation, facilitate neurogenesis, angiogenesis and functional restoration, and save pattern isolation and spatial learning injuries after TBI in animal models [14,45,46]. Herein, the shape, diameter layout, and surface marker proteins of ADSCs-Exo were detected by TEM and Western blot. Moreover, ADSCs-Exo were shown to reduce inflammasome activation in rats after TBI, which contributed to significant sensorimotor functional recovery.
Neurological inflammation is a major molecular and cellular response of the central nervous system (CNS) to trauma. Microglia, the inherent immune cells of the CNS, are a regulator of this response after TBI. Microglial activation initiates the generation of TNF-α and IL-1β/6 [47]. These pro-inflammatory molecules are harmful and activate the auto- and paracrine formation of glutamate from microglia. As such, such activity can directly and neurotoxically impact neurons, synapses and dendrites, depending on the released quantity. LPS-treated primary microglia were used to simulate such neurotoxic microglial behavior [48,49]. The LPS model initiated alterations in microglial activation pro-inflammatory phenotypes. Herein, ADSCs-Exo relieved LPS-caused microglial motion and the secretion of pro-inflammatory factors in vitro. Such results support that microglial inhibition facilitates the treatment effects of ADSCs-Exo on TBI.
Moreover, the secretion of inflammatory cytokines requires inflammasome actions. The NLRP3 inflammasome, consisting of NLRP3, pro-caspase-1 and the adaptor ASC [50], is a significant factor in microglial activation [51] and the best-featured part in microglia. The action of this inflammasome in post-injury neurological inflammation was confirmed in TBI models in vivo and in people with modest/severe TBI. The NLRP3 inflammasome, ASC and caspase-1 mRNA were up-expressed at 6 h and cerebral NLRP3 and caspase-1 protein levels rose at 24 h after a fluid percussion injury [23]. NLRP3, caspase-1 and IL-1β were considerably upregulated in rats at 12 h and 24 h after blast-initiated TBI and in a mouse TBI model [52]. Based on the discovery of the significant function of NLRP3 inflammasome in TBI, it is hypothesized that NLRP3 is a potentially critical target for neuroinflammation modulation and TBI recovery. Actually, NLRP3 silence considerably weakened neuroinflammatory procedures and significantly improved the injured functions in mice [26]. As such, we hypothesize that the anti-TBI function of ADSCs-Exo may also be associated with the start of NLRP3 inflammasome activation. To prove this hypothesis, we examined the inflammatory reaction in a testing model. After the improved sensorimotor functional restoration, ADSCs-Exo blocked the generation of IL-1β/6. ADSCs-Exo also restricted the expression of NLRP3 and caspase-1. The above results infer that ADSCs-Exo may protect rats from TBI by blocking NLRP3-modulated inflammasome stimulation, which then suppresses caspase-1 expression.
ConclusionIn the present study, ADSCs-Exo can improve LPS-caused inflammatory activation by decreasing pro-inflammatory molecules in microglia. Moreover, the neuroprotective role may be ascribed a part to the inhibition on the formation of NLRP3-adjusted inflammasome. This study offers new information about ADSCs-Exo-based therapy and the potential mechanism.
AcknowledgementsThis study was funded by supported by a grant from the Scientific Research Project of the Second People’s Hospital of Wuhu (2020A02, 2020D24, 2019D08) and the Research Project of Teaching Hospital of Wannan Medical College (JXYY202002).
L.T. and Y.X. conceived and designed the experiments. L.T., Y.X., L.W., and J.P. performed experiments. L.W. and L.T. were involved in the analysis and interpretation of data. L.T. and Y.X. were involved in writing and review of the manuscript. L.T. and Y.W. were involved in study supervision.
Conflicts of interestThere are no conflicts of interest.
References 1. Kline AE, Leary JB, Radabaugh HL, Cheng JP, Bondi CO. Combination therapies for neurobehavioral and cognitive recovery after experimental traumatic brain injury: Is more better? Prog Neurobiol 2016; 142:45–67. 2. Crupi R, Cordaro M, Cuzzocrea S, Impellizzeri D. Management of traumatic brain injury: from present to future. Antioxidants (Basel) 2020; 9:297. 3. Cox CS Jr, Baumgartner JE, Harting MT, Worth LL, Walker PA, Shah SK, et al. Autologous bone marrow mononuclear cell therapy for severe traumatic brain injury in children. Neurosurgery 2011; 68:588–600. 4. Li Y, Chopp M. Marrow stromal cell transplantation in stroke and traumatic brain injury. Neurosci Lett 2009; 456:120–123. 5. Nichols JE, Niles JA, DeWitt D, Prough D, Parsley M, Vega S, et al. Neurogenic and neuro-protective potential of a novel subpopulation of peripheral blood-derived CD133+ ABCG2+CXCR4+ mesenchymal stem cells: development of autologous cell-based therapeutics for traumatic brain injury. Stem Cell Res Ther 2013; 4:3. 6. Zhang ZX, Guan LX, Zhang K, Zhang Q, Dai LJ. A combined procedure to deliver autologous mesenchymal stromal cells to patients with traumatic brain injury. Cytotherapy 2008; 10:134–139. 7. Yang Y, Ye Y, Su X, He J, Bai W, He X. MSCs-derived exosomes and neuroinflammation, neurogenesis and therapy of traumatic brain injury. Front Cell Neurosci 2017; 11:55. 8. Chopp M, Li Y. Treatment of neural injury with marrow stromal cells. Lancet Neurol 2002; 1:92–100. 9. Balistreri CR, De Falco E, Bordin A, Maslova O, Koliada A, Vaiserman A. Stem cell therapy: old challenges and new solutions. Mol Biol Rep 2020; 47:3117–3131. 10. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science 2020; 367:eaau6977. 11. Wang MM, Feng YS, Tan ZX, Xing Y, Dong F, Zhang F. The role of exosomes in stroke. Mol Biol Rep 2020; 47:6217–6228. 12. Liu C, Su C. Design strategies and application progress of therapeutic exosomes. Theranostics 2019; 9:1015–1028. 13. Xunian Z, Kalluri R. Biology and therapeutic potential of mesenchymal stem cell-derived exosomes. Cancer Sci 2020; 111:3100–3110. 14. Kim DK, Nishida H, An SY, Shetty AK, Bartosh TJ, Prockop DJ. Chromatographically isolated CD63+CD81+ extracellular vesicles from mesenchymal stromal cells rescue cognitive impairments after TBI. Proc Natl Acad Sci U S A 2016; 113:170–175. 15. de Miranda AS, Zhang CJ, Katsumoto A, Teixeira AL. Hippocampal adult neurogenesis: does the immune system matter? J Neurol Sci 2017; 372:482–495. 16. Devanney NA, Stewart AN, Gensel JC. Microglia and macrophage metabolism in CNS injury and disease: the role of immunometabolism in neurodegeneration and neurotrauma. Exp Neurol 2020; 329:113310. 17. Long X, Yao X, Jiang Q, Yang Y, He X, Tian W, et al. Astrocyte-derived exosomes enriched with miR-873a-5p inhibit neuroinflammation via microglia phenotype modulation after traumatic brain injury. J Neuroinflammation 2020; 17:89. 18. Patel NA, Moss LD, Lee JY, Tajiri N, Acosta S, Hudson C, et al. Long noncoding RNA MALAT1 in exosomes drives regenerative function and modulates inflammation-linked networks following traumatic brain injury. J Neuroinflammation 2018; 15:204. 19. Li Y, Yang YY, Ren JL, Xu F, Chen FM, Li A. Exosomes secreted by stem cells from human exfoliated deciduous teeth contribute to functional recovery after traumatic brain injury by shifting microglia M1/M2 polarization in rats. Stem Cell Res Ther 2017; 8:198. 20. Timmers L, Lim SK, Arslan F, Armstrong JS, Hoefer IE, Doevendans PA, et al. Reduction of myocardial infarct size by human mesenchymal stem cell conditioned medium. Stem Cell Res 2007; 1:129–137. 21. Jaimes Y, Naaldijk Y, Wenk K, Leovsky C, Emmrich F. Mesenchymal stem cell-derived microvesicles modulate lipopolysaccharides-induced inflammatory responses to microglia cells. Stem Cells 2017; 35:812–823. 22. Song H, Zhao C, Yu Z, Li Q, Yan R, Qin Y, et al. UAF1 deubiquitinase complexes facilitate NLRP3 inflammasome activation by promoting NLRP3 expression. Nat Commun 2020; 11:6042. 23. Liu HD, Li W, Chen ZR, Hu YC, Zhang DD, Shen W, et al. Expression of the NLRP3 inflammasome in cerebral cortex after traumatic brain injury in a rat model. Neurochem Res 2013; 38:2072–2083. 24. Ising C, Venegas C, Zhang S, Scheiblich H, Schmidt SV, Vieira-Saecker A, et al. NLRP3 inflammasome activation drives tau pathology. Nature 2019; 575:669–673. 25. Lin C, Chao H, Li Z, Xu X, Liu Y, Bao Z, et al. Omega-3 fatty acids regulate NLRP3 inflammasome activation and prevent behavior deficits after traumatic brain injury. Exp Neurol 2017; 290:115–122. 26. Irrera N, Pizzino G, Calo M, Pallio G, Mannino F, Fama F, et al. Lack of the Nlrp3 inflammasome improves mice recovery following traumatic brain injury. Front Pharmacol 2017; 8:459. 27. Ismael S, Nasoohi S, Ishrat T. MCC950, the selective inhibitor of nucleotide oligomerization domain-like receptor protein-3 inflammasome, protects mice against traumatic brain injury. J Neurotrauma 2018; 35:1294–1303. 28. Si XL, Fang YJ, Li LF, Gu LY, Yin XZ, Jun T, et al. From inflammasome to Parkinson’s disease: does the NLRP3 inflammasome facilitate exosome secretion and exosomal alpha-synuclein transmission in Parkinson’s disease? Exp Neurol 2021; 336:113525. 29. Liang M, Chen X, Wang L, Qin L, Wang H, Sun Z, et al. Cancer-derived exosomal TRIM59 regulates macrophage NLRP3 inflammasome activation to promote lung cancer progression. J Exp Clin Cancer Res 2020; 39:176. 30. Bai L, Li J, Li H, Song J, Zhou Y, Lu R, et al. Renoprotective effects of artemisinin and hydroxychloroquine combination therapy on IgA nephropathy via suppressing NF-kappaB signaling and NLRP3 inflammasome activation by exosomes in rats. Biochem Pharmacol 2019; 169:113619. 31. Yan B, Zhang Y, Liang C, Liu B, Ding F, Wang Y, et al. Stem cell-derived exosomes prevent pyroptosis and repair ischemic muscle injury through a novel exosome/circHIPK3/ FOXO3a pathway. Theranostics 2020; 10:6728–6742. 32. Dai Y, Wang S, Chang S, Ren D, Shali S, Li C, et al. M2 macrophage-derived exosomes carry microRNA-148a to alleviate myocardial ischemia/reperfusion injury via inhibiting TXNIP and the TLR4/NF-kappaB/NLRP3 inflammasome signaling pathway. J Mol Cell Cardiol 2020; 142:65–79. 33. Jha KA, Pentecost M, Lenin R, Gentry J, Klaic L, Del Mar N, et al. TSG-6 in conditioned media from adipose mesenchymal stem cells protects against visual deficits in mild traumatic brain injury model through neurovascular modulation. Stem Cell Res Ther 2019; 10:318. 34. Xu C, Diao YF, Wang J, Liang J, Xu HH, Zhao ML, et al. Intravenously infusing the secretome of adipose-derived mesenchymal stem cells ameliorates neuroinflammation and neurological functioning after traumatic brain injury. Stem Cells Dev 2020; 29:222–234. 35. Santos J, Milthorpe BK, Padula MP. Correction: Santos, J., et al. Proteomic analysis of cyclic ketamine compounds ability to induce neural differentiation in human adult mesenchymal stem cells. International Journal of Molecular Sciences 2019, 20, 523. Int J Mol Sci 2019; 20:3542. 36. Chen H, Li Z, Lin M, Lv X, Wang J, Wei Q, et al. MicroRNA-124-3p affects myogenic differentiation of adipose-derived stem cells by targeting Caveolin-1 during pelvic floor dysfunction in Sprague Dawley rats. Ann Transl Med 2021; 9:161. 37. Radabaugh HL, Carlson LJ, O’Neil DA, LaPorte MJ, Monaco CM, Cheng JP, et al. Abbreviated environmental enrichment confers neurobehavioral, cognitive, and histological benefits in brain-injured female rats. Exp Neurol 2016; 286:61–68. 38. Beery AK, Zucker I. Sex bias in neuroscience and biomedical research. Neurosci Biobehav Rev 2011; 35:565–572. 39. Gierloff M, Petersen L, Oberg HH, Quabius ES, Wiltfang J, Acil Y. Adipogenic differentiation potential of rat adipose tissue-derived subpopulations of stromal cells. J Plast Reconstr Aesthet Surg 2014; 67:1427–1435. 40. Deierborg T. Preparation of primary microglia cultures from postnatal mouse and rat brains. Methods Mol Biol 2013; 1041:25–31. 41. Chen J, Sanberg PR, Li Y, Wang L, Lu M, Willing AE, et al. Intravenous administration of human umbilical cord blood reduces behavioral deficits after stroke in rats. Stroke 2001; 32:2682–2688. 42. Baskin YK, Dietrich WD, Green EJ. Two effective behavioral tasks for evaluating sensorimotor dysfunction following traumatic brain injury in mice. J Neurosci Methods 2003; 129:87–93. 43. Kim YJ, Lee KS, Joe SG, Kim JG. Incidence and quantitative analysis of dissociated optic nerve fiber layer appearance: real loss of retinal nerve fiber layer? Eur J Ophthalmol 2018; 28:317–323. 44. Jeppesen DK, Fenix AM, Franklin JL, Higginbotham JN, Zhang Q, Zimmerman LJ, et al. Reassessment of Exosome Composition. Cell 2019; 177:428–445.e18. 45. Williams AM, Bhatti UF, Brown JF, Biesterveld BE, Kathawate RG, Graham NJ, et al. Early single-dose treatment with exosomes provides neuroprotection and improves blood-brain barrier integrity in swine model of traumatic brain injury and hemorrhagic shock. J Trauma Acute Care Surg 2020; 88:207–218. 46. Chen Y, Li J, Ma B, Li N, Wang S, Sun Z, et al. MSC-derived exosomes promote recovery from traumatic brain injury via microglia/macrophages in rat. Aging (Albany NY) 2020; 12:18274–18296. 47. Ransohoff RM, Brown MA. Innate immunity in the central nervous system. J Clin Invest 2012; 122:1164–1171. 48. Fenn AM, Gensel JC, Huang Y, Popovich PG, Lifshitz J, Godbout JP. Immune activation promotes depression 1 month after diffuse brain injury: a role for primed microglia. Biol Psychiatry 2014; 76:575–584. 49. Mao SS, Hua R, Zhao XP, Qin X, Sun ZQ, Zhang Y, et al. Exogenous administration of PACAP alleviates traumatic brain injury in rats through a mechanism involving the TLR4/MyD88/NF-kappaB pathway. J Neurotrauma 2012; 29:1941–1959. 50. Mamik MK, Power C. Inflammasomes in neurological diseases: emerging pathogenic and therapeutic concepts. Brain 2017; 140:2273–2285. 51. He W, Long T, Pan Q, Zhang S, Zhang Y, Zhang D, et al. Microglial NLRP3 inflammasome activation mediates IL-1beta release and contributes to central sensitization in a recurrent nitroglycerin-induced migraine model. J Neuroinflammation 2019; 16:78. 52. Ma J, Xiao W, Wang J, Wu J, Ren J, Hou J, et al. Propofol inhibits NLRP3 inflammasome and attenuates blast-induced traumatic brain injury in rats. Inflammation 2016; 39:2094–2103.
Comments (0)