
Remember me

In this translational study, we combined assessments of tau PET, in vitro autoradiography, quantitative tau immunohistochemistry, and cellular tracer uptake using a 4R-tauopathy mouse model and human samples consisting of patients with 4R-tauopathies and disease controls.
Small animal experimentsAll small animal experiments were approved by the local animal care committee of the Government of Upper Bavaria (Regierung Oberbayern, approval number: ROB-55.2-2532.Vet_02-15-210, ROB-55.2-2532.Vet_02-19-26). The experiments were overseen by a veterinarian and conducted in compliance with the ARRIVE guidelines and in accordance with the U.K. Animals (Scientific Procedures) Act, 1986 and associated guidelines, EU Directive 2010/63/EU for animal experiments. The animals were housed in a temperature- and humidity-controlled environment with a 12 h light‒dark cycle and free access to food (Ssniff Spezialdiäten GmbH, Soest, Germany) and water. Anesthesia was induced before [18F]PI-2620 application and maintained during the PET and MR scans, with 1.5% isoflurane delivered via a mask at 3.5 L/min. All procedures were performed at the Department of Nuclear Medicine, Ludwig Maximilian University (LMU) Hospital, Munich. First, we conducted a longitudinal [18F]PI-2620 PET/MRI study in a 4R-tau mouse model (PS19) and age-matched wild-type mice (n = 10 each, all female) using regional tau PET signals and volumetric measures as endpoints. PS19 transgenic mice express mutated human microtubule-associated protein tau (MAPT) under the control of the mouse prion protein (Prnp) promoter. This transgene encompasses the P301S mutation linked to the disease and contains four microtubule-binding domains alongside an N-terminal insert (4R/1N) [57]. Next, we performed immunohistochemistry in a subset of these PS19 and wild-type mice (n = 4 each) to characterize the regional tau abundance and cellular contributions to tau pathology. Cell sorting after tau radiotracer injection was applied in another subset of PS19 and wild-type mice (n = 5 each) to determine the cellular origin of the tau PET signals.
Human analysesA key experiment of the study consisted of a correlation analysis between regional [18F]PI-2620 tau PET signals and the abundance of fibrillary tau pathology in autopsy samples from patients with definite PSP (n = 6) and disease controls (n = 2, amyotrophic lateral sclerosis, TDP-43-positive frontotemporal lobe degeneration). In this sample, we performed a quantitative correlation analysis between tau PET signals, autoradiography, and the abundance of AT8-positive tau pathology. An additional autopsy sample from deceased patients with PSP presenting with limited copathology (n = 16) was used to determine the contributions of tau-positive neurons and tau-positive astrocytes to the [18F]PI-2620 autoradiography signals. To this end, the abundance of AT8-positive tau pathology in subfields (≥ 8) of the frontal cortex and basal ganglia sections was differentiated between neurons and astrocytes using a data-driven tissue classifier. An orienting a priori sample size calculation suggested a minimum of n = 6 subjects or subfields to achieve a statistical power of 0.8 at α = 0.05 for the detection of a meaningful explanation of variance (β ≥ 0.3; residual variance of 0.15) by two predictors (i.e., neuronal and astrocytic tau abundances). The tissue samples from all the autopsy cases investigated were provided by Neurobiobank Munich, LMU Munich. They were collected according to the guidelines of the local ethics committee, and the usage of the material for this project was additionally approved (application number 19-244). Finally, the combined findings were used to evaluate the emerging gray matter/white matter boundary target region for a tau PET assessment of 4R-tau pathology in the cortex of patients with PSP (n = 17) compared with controls (n = 9). All patients and controls who underwent in vivo PET imaging provided informed written consent. The study was conducted in accordance with the principles of the Declaration of Helsinki, and approval was obtained from the local ethics committee (application numbers 17-569 and 19-022).
Small animal PET/MRI imagingPET/MRI acquisitionAll rodent PET procedures followed an established standardized protocol for radiochemistry, acquisition times, and postprocessing using a PET/MRI system. All the mice were scanned with a 3 T Mediso nanoScan PET/MR scanner (Mediso Ltd., Hungary) with a triple-mouse imaging chamber. Two 2-min anatomical T1 MR scans were performed prior to tracer injection (head receiver coil, matrix size 96 × 96 × 22, voxel size 0.24 × 0.24 × 0.80 mm3, repetition time 677 ms, echo time 28.56 ms, and flip angle 90°). The injected dose of [18F]PI-2620 delivered in 200 µl saline via intravenous injection was 12.7 ± 2.1 MBq. PET emission was recorded in a dynamic 0–60 min window. The frames used were 6 × 10, 2 × 30, 3 × 60, 5 × 120, 5 × 300, and 5 × 600. The list-mode data within the 400–600 keV energy window were reconstructed using a 3D iterative algorithm (Tera-Tomo 3D, Mediso Ltd., Hungary) with the following parameters: matrix size of 55 × 62 × 187 mm3, voxel size of 0.3 × 0.3 × 0.3 mm3, 8 iterations, and 6 subsets. Decay, random, and attenuation corrections were applied. The T1 image was used to create a body–air material map for attenuation correction. We longitudinally studied PS19 (n = 10) and age-matched wild-type mice (n = 10, WT; C57BL6) at 5.9, 7.7, 10.2, and 12.4 months of age. The sample size was selected based on the assumption of detecting a 10% difference between genotypes at the latest time point with a power of 0.8, applying an α of 0.05. No randomization was used to allocate the experimental units due to the absence of any intervention. No dropouts were registered; hence, all the mice were included in the subsequent analysis. Blinding was not applied during the scanning process, but it was implemented during image analysis, where an automatic coregistration step guaranteed reader independence [38].
PET/MRI analysesThe normalization of the PET data was performed by calculating the volume of distribution (VT) images obtained from the full dynamic scan, as described previously for different tracers [6, 56]. Briefly, we generated VT images with an image-derived input function using the methodology described by Logan et al. implemented in PMOD. The plasma curve was obtained from a standardized bilateral VOI placed in the left ventricle. A maximum error of 10% and a VT threshold of 0 were selected for modeling the full dynamic imaging data. Furthermore, 20–40 min static [18F]PI-2620 images were analyzed as a readout matching the scRadiotracing normalization. We applied the striatum as a reference tissue to decrease the variability at the individual subject level and calculated the VT ratio and standardized uptake value ratio (SUVR) for images. The reference tissue was validated by analyzing VT images from PS19 and WT mice, which confirmed that no differences in VT in the striatum (8.4 mm3) were observed between genotypes. Predefined volumes of interest were delineated by spheres in the brainstem (4.2 mm3) and the entorhinal cortex (2.8 mm3), guided by regions of the Mirrione atlas but eroded to avoid the spill-in of adjacent brain structures (Supplemental Fig. 1). These target regions served for the extraction of PET values for all the mice.
The MRI volumetric analysis was performed in a blinded manner on coronal sections by manual delineation of the cerebellum, the brainstem, and the striatum (each in three adjacent planes) using PMOD (Supplemental Fig. 1). We conducted a test–retest procedure to ensure the reliability of MRI segmentation, which displayed a high congruency (r) of > 0.9 across 10 test cases. The cerebellum and brainstem were considered as a combined hindbrain region.
scRadiotracingMouse brain dissociationFive PS19 and five WT mice underwent scRadiotracing [7, 8, 56] immediately after the tau PET scan. An Adult Brain Dissociation Kit (mouse and rat) (Miltenyi Biotec, 130-107-677) was used for brain dissociation according to the manufacturer's instructions. Adult mouse brains were dissected, briefly washed with phosphate-buffered saline (PBS), cut into eight pieces, and dissociated with enzyme mixtures 1 and 2 using a gentleMACS™ Octo Dissociator (Miltenyi Biotec, 130-096-427). The dissociated cell suspension was applied to a prewet 100 µm cell strainer (Falcon, 352,360). The cell pellet was resuspended in cold PBS and cold debris removal solution. Cold PBS was gently overlaid on the cell suspension. The mixture was centrifuged at 4 °C and 3000×g for 10 min with acceleration and deceleration set at 5. The two top phases were removed entirely. The cell pellets were collected and resuspended in 1 ml of cold red blood cell removal solution, followed by 10 min of incubation. The cell pellets were collected for astrocyte and subsequent neuronal isolation via magnetic activated cell sorting (MACS) [7, 8, 56].
Isolation of astrocytesAn Adult Brain Dissociation Kit for mouse and rat (Miltenyi Biotec, 130-107-677) was used according to the manufacturer's instructions. The prepared cell pellets were resuspended in 80 µl of AstroMACS separation buffer (Miltenyi Biotec, 130-117-336) per 107 total cells. Then, 10 μL of FcR blocking reagent was added, and the mixture was incubated for 10 min in the dark at 4 °C. Next, 10 μL of Anti-ACSA-2 MicroBeads was added, and the mixture was incubated for 15 min in the dark at 4 °C. The cells were washed by adding 1 mL of AstroMACS separation buffer and centrifuged at 300×g for 5 min. The cell pellets were resuspended in 500 μL of AstroMACS separation buffer. The prewet MS columns (Miltenyi Biotec, 130-042-201) were placed in an OctoMACS Separator (Miltenyi Biotec, 130-042-109). The cell suspensions were applied onto the column, followed by washes with 3 × 500 µL of AstroMACS separation buffer. The flow-through was collected and contained nonastrocytic cells as an astrocyte-depleted fraction. The columns were removed from the magnetic field, and the astrocytes were flushed out using 3 ml of AstroMACS separation buffer.
Isolation of neuronsA Neuron Isolation Kit, mouse (Miltenyi Biotec, 130-115-390) was used as previously reported [20], according to the manufacturer's instructions. The astrocyte-depleted cell pellets were resuspended in 80 µl of PBS–0.5% bovine serum albumin (BSA) buffer per 107 total cells. Twenty microliters of nonneuronal cell biotin–antibody cocktail was added, and the mixture was incubated for 5 min in the dark at 4 °C. The cells were washed and centrifuged at 300×g for 5 min. The cell pellets were again resuspended in 80 μL of PBS–0.5% BSA buffer per 107 total cells. Next, 20 μL of anti-biotin microbeads was added, and the mixture was incubated for 10 min in the dark at 4 °C. The volume was adjusted to 500 µl per 107 total cells with PBS–0.5% BSA buffer, and then, magnetic separation was performed. The prewet LS columns (Miltenyi Biotec, 130-042-401) were placed in a QuadroMACS™ Separator (Miltenyi Biotec, 130-090-976). The cell suspensions were applied onto the columns. The columns were washed with 2 × 1 ml of PBS–0.5% BSA buffer. The flow-throughs containing the unlabeled cells were collected as the neuron-enriched fractions. The columns were removed from the magnetic field, and the nonneuronal cells were flushed out with 3 ml of PBS–0.5% BSA buffer [56].
Gamma emission, flow cytometry, and calculation of single-cell tracer uptakeThe radioactivity concentrations of the cell pellets were measured with a highly sensitive gamma counter (Hidex AMG Automatic Gamma Counter, Mainz, Germany) are reported relative to the activity in the whole brain, with decay correction to the time of tracer injection for the final activity calculations.
Flow cytometry staining was performed at 4 °C. After the gamma emission measurement, the cell suspension was centrifuged at 400×g for 5 min, and the supernatant was aspirated completely. The cell pellet was then resuspended in 100 µl of cold D-PBS containing fluorochrome-conjugated antibodies recognizing mouse CD11b and ACSA-2 (Miltenyi Biotec, 130-113-810 and 130-116-247) at a 1:100 dilution and incubated for 10 min at 4 °C in the dark. The samples were washed with 2 ml of D-PBS and centrifuged for 5 min at 400×g. Finally, the cell pellets were resuspended in 500 μl of D-PBS, and the samples were immediately used for flow cytometry with an MACSQuant® Analyzer as a quality control for MACS. Absolute cell numbers were acquired for all the samples. The purity of the astrocyte-enriched cell pellet was determined via ACSA-2 staining. For the assessment of purity within the neuron-enriched fraction, the proportion of remaining CD11b- and ACSA-2-positive cells was determined and subtracted from the total number of cells within the pellet. The CD11b-/ACSA-2-negative fraction was considered neurons and validated using CD90.1 as a neuronal marker for C57BL6 mice (consistently > 85%).
The measured radioactivity (Bq) of the cell pellets was divided by the specific cell number in the pellet, resulting in the calculated radioactivity per cell. The sufficient sensitivity of a single readout was determined as a ≥ twofold ratio between the cell pellet radioactivity and the background measurement, with a total procedure duration of 6–7 h from tracer injection to radioactivity measurement in the enriched cell pellets. The radioactivity per cell was normalized to the injected radioactivity and body weight (%ID*BW).
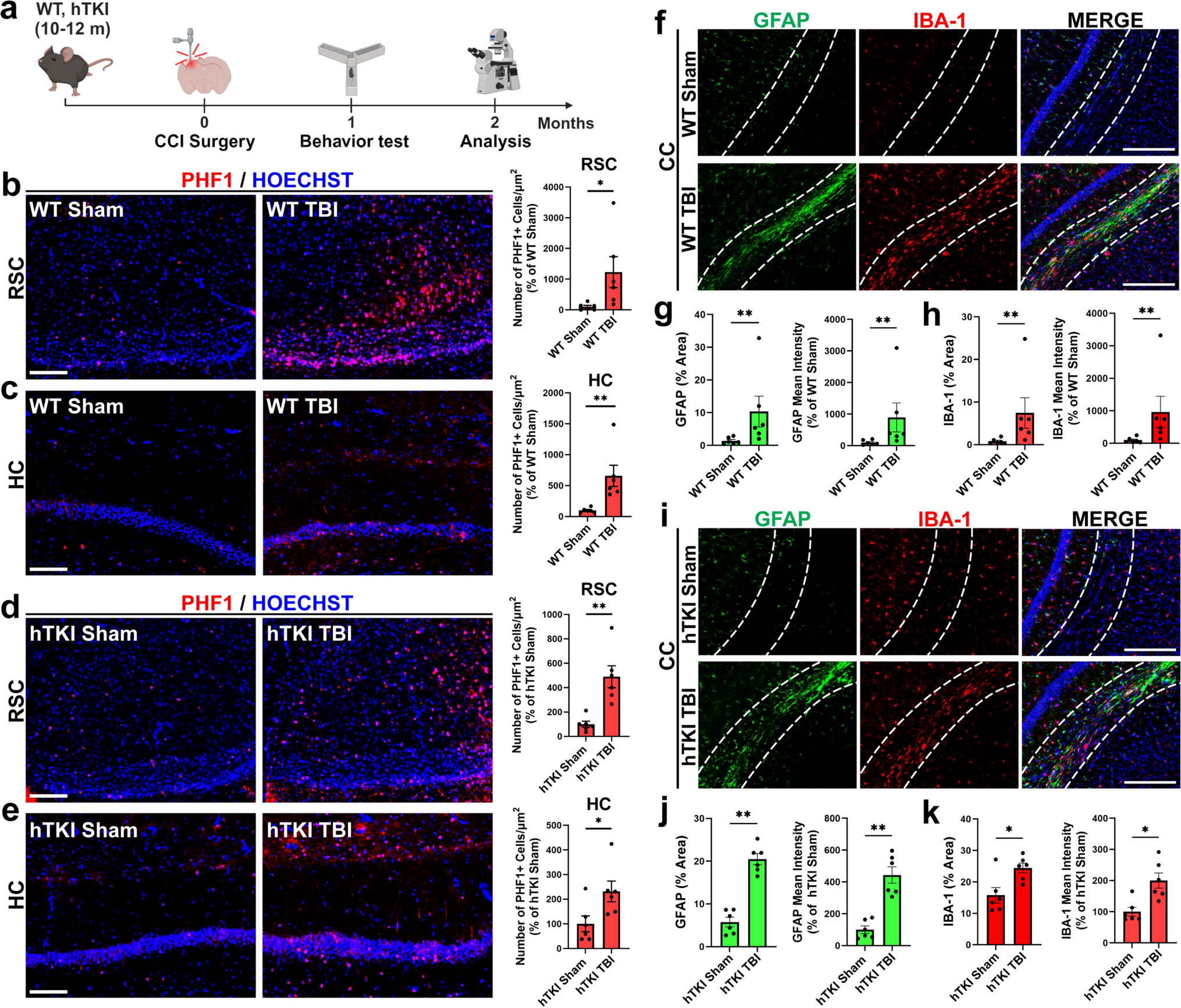
Small animal immunohistochemistryAn additional number of four female PS19 mice and four wild-type mice were used for immunohistochemistry, as the same brains could not be used for both scRadiotracing analysis and immunohistochemical staining. Fifty-micron-thick slices were cut in the sagittal plane using a vibratome (VT1200S, Leica Biosystems). Slices were treated with blocking solution (10% normal goat serum and 10% donkey serum in 0.3% Triton and PBS to a total volume of at least 200 μl per well/slice) for 3 h at RT. The following primary antibodies were used: chicken anti-GFAP (1:500; ab5541; Merck Millipore, Darmstadt, Germany), mouse anti-AT8 (1:1000; ab5541; Merck Millipore, Darmstadt, Germany), and rabbit anti-MAP2 (1:500) diluted in blocking solution (5% normal goat serum and 5% donkey serum in 0.3% Triton and PBS to a total volume of at least 200 μl per well/slice). The antibodies were applied to the slices and subsequently incubated for ~ 48 h at 4 °C on a horizontal shaker. The following secondary antibodies were used: goat anti-rabbit Alexa Fluor 488 (1:500), goat anti-chicken Alexa Fluor 555 (1:500), and goat anti-mouse Alexa Fluor 647 (1:500) diluted in PBS. Slices were incubated for 2–3 h at RT on a horizontal shaker in the dark. After 3 × 10 min washes with PBS, the slices were mounted and cover slipped with fluorescence mounting medium containing DAPI (Dako, Santa Clara, USA).
Three-dimensional images were acquired with an Apotome microscope (Zeiss Oberkochen, Germany) using 10 × and 40 × objectives. The analysis programs Zeiss blue and ImageJ were used for quantification. Z-stack images (10 μm) were acquired with a 10 × objective. Each signal (AT8 and GFAP) was quantified as the % area of the entire scanning frame.
Single optical sections were acquired using a 40 × objective, and the AT8 signal was analyzed, because the perceptual signals were MAP2- and GFAP-positive. To this end, we created a mask of the AT8 signal and transferred it into GFAP-positive astrocytes and MAP2-positive neuronal structures. After local brightness/contrast adjustments and background subtraction, we set a fixed threshold and calculated the AT8 area (%) inside the mask of the GFAP-positive astrocytes and MAP-positive neurons.
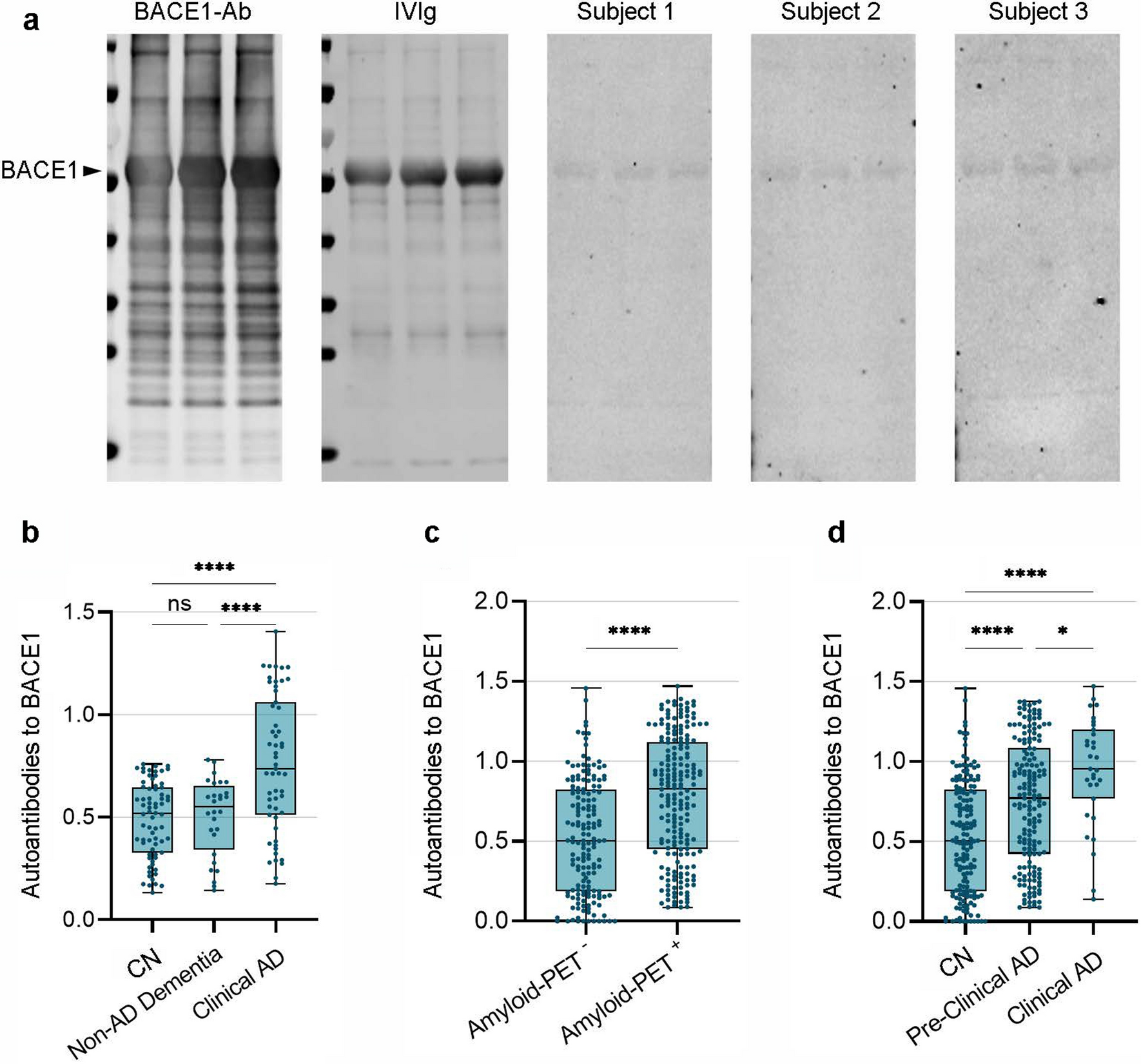
Human postmortem samplesHuman samplesFor PET of autopsy samples, we included all patients who underwent [18F]PI-2620 tau PET prior to death, donated their brain to the Munich Brain Bank, and underwent a tissue workup by 31 March 2024 (n = 8; Supplemental Table 1). The formalin-fixed and paraffin-embedded tissue blocks of one hemisphere were used for AT8 and autoradiography analyses. The tissue from the medial frontal gyrus and basal ganglia, including the globus pallidus, was available for seven patients with definite PSP, one patient with FTLD-TDP, and one patient with FTLD/MND-TDP. For in-depth analyses of the origins of the cellular and structural radiotracer signals, we selected samples with limited α-synuclein, TDP-43, and FUS pathology from the Munich Brain Bank. Limited β-amyloid pathology was tolerated, resulting in a total sample size of n = 16 (Supplemental Table 2). Intact medial frontal gyrus tissue was available for fourteen patients, and intact basal ganglia tissue, including the globus pallidus, was available for seven patients. We conducted [18F]PI-2620 autoradiography and AT8 immunohistochemistry on postmortem brain tissues from n = 4 patients with clinically diagnosed Parkinson’s disease (PD) to examine the specificity of tracer binding (Supplemental Table 3).
ImmunohistochemistryImmunohistochemistry was performed on 4 µm-thick sections of formalin-fixed and paraffin-embedded tissue using standard techniques. Immunohistochemical tau staining was performed semiautomatically using a BenchMark device (Ventana, now Hoffmann-LaRoche, Basel, Switzerland) with a mouse monoclonal AT8 antibody raised against hyperphosphorylated tau (Ser202/Thr205, 1:200, Invitrogen/Thermo Fisher, Carlsbad, CA, USA), as well as with the mouse monoclonal isoform-specific tau antibodies RD3 (8E6/C11) and RD4 (1E1/A6), on adjacent sections of those used for ARG. The immunostained sections were digitized at 20 × magnification with a Mirax Midi scanner (Zeiss, Carl Zeiss MicroImaging GmbH, Jena, Germany). For frontal cortex (medial frontal gyrus) and globus pallidus analyses, 8–12 regions of interest (subfields) were drawn manually per section, and the AT8-positive tau load (%) was quantified using ZEN 3.4 blue edition software (Zeiss, Jena, Germany).
Similar to previous approaches, i.e., by Rittman et al. [40], we aimed to subdivide the AT8-positive tau load into different underlying cell types. Therefore, we used semiautomated object characterization and recognition to differentiate neurofibrillary tangles (NFTs), coiled bodies (CBs), and tufted astrocytes (TAs) based on several parameters of morphological characteristics. Blinding was ensured by randomizing and renaming the digital images, effectively eliminating any potential bias associated with the sample origin. Single NFT, CB, and TA (n = 15–20 objects per slice) were manually selected to define object thresholds, including the size (area), diameter, ellipse axis, perimeter, intensity, grade of circularity, roundness, and compactness. Specific masks were generated for positive NFTs, CBs, and TAs and were uniformly applied to all sections (Supplemental Fig. 2). This combination of blinding, randomization, and consistent mask application ensured reproducibility and allowed for the precise quantification of AT8 immunoreactivity across the entire sample set. Due to substantial overlap of object characteristics, we subsequently defined NFT and CB as a combined group of AT8-positive cells with high density. Notably, tau fragments (TFs) were partially included in the TA channel, resulting in two analysis channels (NFT/CB and TA/TF). This finding was substantiated by the correlation analysis between AT8 positivity and autoradiography signals in single subjects, which revealed similar associations of neuron- and oligodendrocyte-enriched regions with autoradiography signals. The final segmentation resulted in NFT/CB AT8-area-%, TA/TF AT8-area% and intensities within 8–12 subfields per analyzed section.
For the correlation analysis between in vivo PET imaging data and the tau load at autopsy, composite regions of interest in the medial frontal gyrus and in the globus pallidus (internal and external parts) were used.
AutoradiographyFor direct comparison with the autoradiography signal, tau immunostaining of formalin-fixed and paraffin-embedded tissue blocks from 16 PSP patients and 4 PD patients and three brain regions (frontal cortex, putamen, and pallidum) was performed. For each patient and brain region, autoradiography with [18F]PI-2620 was performed on ≥ 4 sections, as described previously [55]. Briefly, the sections were incubated for 45 min (21.6 μCi/ml after dilution to a volume of 50 ml with phosphate-buffered saline solution, pH 7.4, specific activity of 480 ± 90 GBq/μmol), washed, dried, placed on imaging plates for 12 h, and scanned at 25.0 µm resolution. Regions of interest were drawn on each sample using the AT8 staining of the adjacent section, thus serving to anatomically define subfields in the frontal cortex (gray matter and white matter). An AT8-negative region in the white matter was used as the reference region, and the ratios between the subfield target regions and the reference region were calculated. Each subfield region was labeled with a cortical or gray matter/white matter boundary. Binding ratios were correlated with a semiquantitative AT8 assessment using Pearson’s correlation coefficient after testing for normality and subjected to a regression analysis (neuronal vs. astrocytic tau).
Human PET imaging and analysisTau PET image acquisition and preprocessing[18F]PI-2620 was synthesized as previously described [47]. The injected dose ranged between 156 and 223 MBq and was applied as a slow (10 s) intravenous bolus injection. Positron emission tomography (PET) imaging was performed in a fully dynamic setting (scan duration: 0–60 min postinjection) using a Siemens Biograph True point 64 PET/CT system or a Siemens mCT system (Siemens, Erlangen, Germany). The dynamic brain PET data were acquired in three-dimensional list mode over 60 min and reconstructed into a 336 × 336 × 109 matrix (voxel size: 1.02 × 1.02 × 2.03 mm3) using the built-in ordered subset expectation maximization (OSEM) algorithm with 4 iterations, 21 subsets, and a 5 mm Gaussian filter on the Siemens Biograph and with 5 iterations, 24 subsets, and a 5 mm Gaussian filter on the Siemens mCT. Low-dose CT served for attenuation correction. Frame binning was standardized to 12 × 5 s, 6 × 10 s, 3 × 20 s, 7 × 60 s, 4 × 300 s, and 3 × 600 s. Image-derived input functions were generated by manual and automated extraction of the PET standardized uptake value (SUV) from the carotid artery over a 60-min dynamic PET scan.
Via manual extraction, the blood activity concentration in the bilateral carotid artery was detected in early frames of the dynamic PET images, and spheres with a diameter of 5.0 mm were placed as volumes of interest (VOIs) in the pars cervicalis of the internal carotid artery prior to entering the pars petrosal using PMOD version 4.2 (PMOD Technologies, Zürich, Switzerland). The activity concentration over time was calculated from the average values of the VOI.
Tau PET quantificationVolume distribution (VT) images were calculated with the IDIFs using Logan plots [30], which assume that the data become linear after an equilibration time t*. t* was fitted based on the maximum error criterion, which indicates the maximum relative error between the linear regression and the Logan-transformed measurements in the segment starting from t*. The maximum error was set to 10%. The percentage of masked pixels was set to 0%. The putamen, which was defined by manual placement of a VOI (sphere with a diameter of 10 mm), served as the tissue region.
All VT images were transformed to MNI space via 20–40 min coregistration using the established [18F]PI-2620 PET template [18]. The automated brain normalization settings in PMOD included nonlinear warping, 8 mm input smoothing, equal modality, 16 iterations, a frequency cutoff of 3, regularization of 1.0, and no thresholding.
For the PET-to-autopsy correlation analysis, VT ratios (temporal white matter reference region [10]) were obtained in the medial frontal gyrus and in the globus pallidus as PSP target regions (see immunohistochemistry), which were predefined by the atlas of the basal ganglia [25], the Brainnetome atlas [17], and the Hammers atlas [21]. The rationale was to employ a matched quantification strategy between PET and autoradiography.
GM/WM target regionAll patients and controls used for this dedicated analysis underwent T1-weighted structural MRI on a 3 T Siemens Magnetom PRISMA or SKYRA Scanner and [18F]PI-2620 tau PET in a fully dynamic setting (0–60 min postinjection) using pre-established standard PET scanning parameters [46]. Our in-house [18F]PI-2620 synthesis and quality control pipeline has been described in detail previously [12, 19]. Dynamic [18F]PI-2620 tau PET images were acquired on a Siemens Biograph True point 64 PET/CT (Siemens, Erlangen, Germany) or a Siemens mCT scanner (Siemens, Erlangen, Germany) in 3D list mode over 60 min together with low-dose CT for attenuation correction. For dynamic PET image acquisition, we reconstructed late-phase tau PET images at 20–40 min p.i., which were summarized into a single frame after motion correction [9, 24, 46, 53].
All images were screened for artifacts before preprocessing. T1-weighted structural MR scans were bias-corrected and segmented into tissue types using the CAT12 toolbox (https://neuro-jena.github.io/cat12-help/). PET images were linearly coregistered to the corresponding T1 MRI data, and the intensity was normalized using a pre-established inferior cerebellar reference region [19]. Using T1 MRI data, surface reconstruction was performed using the CAT12-based cortical thickness pipeline. Surface reconstruction was performed for the GM/WM boundary and then systematically shifted to the underlying white matter, as well as to the GM/CSF boundary. Using these systematically shifted surfaces, we extracted PI-2620 tau PET SUVRs for 200 regions of the cortical Schaefer atlas to systematically determine the tau PET signal from the GM/CSF toward the GM/WM boundary and below.
StatisticsGraphPad Prism (V10, GraphPad Software, US) and SPSS (V27, IBM, US) were used for the statistical analyses.
Mouse PET/MRIMixed linear models (Graph Pad Prism) were used to test for age × genotype effects on PS19 and WT mice, including tau PET binding and MRI volumes, between 6 and 12 months of age as indices of interest. Pearson’s correlation coefficient was calculated between brainstem tau PET binding and the brainstem volume at all investigated time points (separately for PS19 and WT mice). As a supporting analysis, we correlated tau PET binding and immunohistochemistry in the frontal cortex (n = 4) and the hippocampus (n = 3), which are brain regions with well-documented tau protein immunoreactivity [3] and limited spill-over from adjacent tissue, to ensure reliable values at the single-mouse-level of individual PS19 mice. One hippocampal section was excluded due to limited quality of the tissue material. A sample size calculation was performed (G*Power) to determine the minimal age of detectability of tau PET binding and MRI volume with cohorts of n = 12 PS19 mice and n = 12 WT mice (power = 0.8, α = 0.05; Supplemental Fig. 3). No corrections for multiple comparisons were applied in the small animal PET experiments, as only a limited number of target regions were selected. This approach resulted in a low Type I error rate (α), with moderate sample sizes, thereby adhering to the 3Rs (Reduction) principles of animal welfare.
Mouse immunohistochemistryRegional coverage of AT8 and GFAP staining was compared between PS19 and WT mice with an unpaired Student’s t test. The colocalization of AT8 with neurons (MAP2 +) and astrocytes (GFAP +) was compared using an unpaired Student’s t test.
Mouse scRadiotracingRadiotracer uptake per neuron and astrocyte was compared between PS19 and WT mice using an unpaired Student’s t test. Furthermore, in PS19 and WT mice, radiotracer uptake was compared between neurons and astrocytes. Pearson’s correlation coefficients were calculated between the radioactivity per cell and the tau PET signals for the combined data from the PS19 and WT mice, as well as for the subset of PS19 mice. For the whole-brain voxelwise correlation analysis between cellular tracer uptake (n = 5 PS19, n = 5 WT), statistical parametric mapping (SPM) was performed using SPM12 routines (Wellcome Department of Cognitive Neurology, London, UK) implemented in MATLAB (version 2016). Individual SUVR images were subjected to linear regression analysis with cellular tracer uptake in neurons or astrocytes (%ID*BW) as a vector in the pooled cohort of PS19 and WT mice (threshold: p < 0.005 uncorrected, k > 20 voxels). Increases in the tau PET signals in each of the five PS19 mice that underwent scRadiotracing were compared with the average tau PET signal in the WT mice. The average tau PET signal in the WT mice was considered an unspecific background signal, as the WT mice did not show any tau accumulation in the brain. As PET signals need to be recognized as a product of cellular tracer uptake and cell type abundance in the brain, we used extrapolation to estimate the total contributions of neurons and astrocytes to increases in tau-related PET signals in PS19 mice. We multiplied the individual cellular tracer uptake (astrocytes and neurons) of each mouse with published cell numbers (i.e., 71*10e6 neurons and 21*10e6 astrocytes) as a surrogate for the cell type abundance in the brains of PS19 mice [22]
$$\begin 7110}^ \left( }}}}}}Mouse\# 1 + \frac}}}}}}Mouse\# 2 + \frac}}}}}}Mouse\# 3 + \frac}}}}}}Mouse\# 4 + \frac}}}}}}Mouse\# 5} \right) \hfill \\ + 2110}^ \left( }}}}}}Mouse\# 1 + \frac}}}}}}Mouse\# 2 + \frac}}}}}}Mouse\# 3 + \frac}}}}}}Mouse\# 4 + \frac}}}}}}Mouse\# 5} \right) \hfill \\ = }_}} \left( } \right) + }_}} \left( } \right) \ldots + }_}} \left( } \right). \hfill \\ \end$$
We used the sum of all 5 mice for comparison instead of single mice to minimize confounding factors caused by, e.g., distinct cell type abundances in the brains of individual mice compared with those reported in the literature or the methodological variance of scRadiotracing. A paired t test was used to compare PET radioactivity and the extrapolated radioactivity of single cells in the cohort of five PS19 mice.
Human PET-to-autopsy correlationPartial correlation coefficients were calculated for the multimodal correlation between tau abundance via immunohistochemistry, autoradiography ratios, and tau PET signals, accounting the globus pallidus and frontal cortex as cofactors across samples.
Human autoradiographyNeuronal and astroglial tau abundances were compared with paired t tests. The Kolmogorov‒Smirnov test confirmed the normality of the residuals. Neuronal and astroglial tau abundances in subfields of the frontal cortex were correlated with autoradiography binding ratios in corresponding regions (total n = 129; Supplemental Fig. 4). Additionally, a linear regression analysis was performed with neuronal and astroglial tau abundances as predictors and autoradiography binding ratios as the outcome variable. At the individual level, the correlation between neuronal and astroglial tau abundances and autoradiography binding ratios was determined in 8–12 subfields per subject. These individual correlations were analyzed as a function of overall tau abundance (separately for neurons and astroglia). In basal ganglia regions, the AT8 signal intensity and AT8 occupancy were correlated with autoradiography binding ratios.
Human PET target regionDifferences in the SUVRs of n = 5 different layers (GM/CSF boundary, GM toward CSF, GM toward WM, GM/WM boundary, and below the GM/WM boundary) were compared using a repeated-measures ANOVA, accounting for within-subject variability. Post hoc pairwise comparisons were performed using Tukey-adjusted estimated marginal means to identify specific differences between layers.
Comments (0)