
Remember me
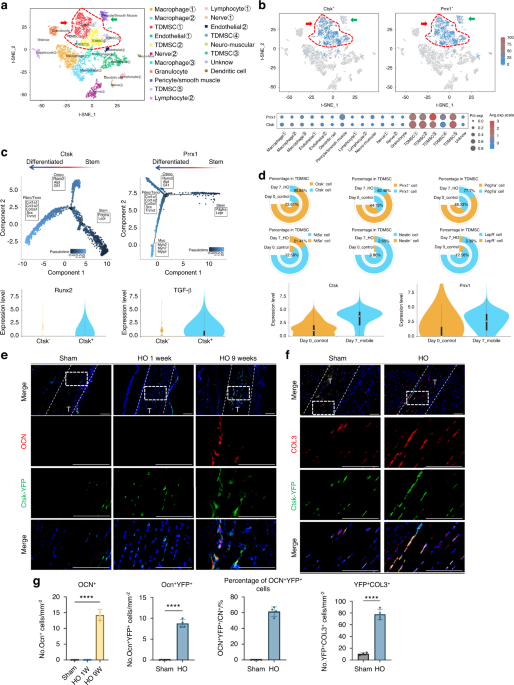
To identify the phenotype of aberrantly differentiated MSCs after injury, we performed unbiased single-cell sequencing data analysis of the entire local tissue after Achilles tendon injury modeling (GEO: GSE150995). We classified mesenchymal cells with higher expression of Prrx1 and Pdgfrα which have stronger stem cell characteristics, as TDMSCs (Fig. S1A). The results of the cell fractionation t-SNE plots for single-cell analysis are displayed in Fig. 1a. Compared with the uninjured area, the number of TDMSCs in the injured area significantly increased after injury-induced heterotopic ossification (HO) (Fig. S2A,B). We found that Ctsk was specifically expressed in mesenchymal stem cells in the Achilles tendon area than was the classical mesenchymal stem cell marker Prrx1, the expression of which was also detected in other types of cells, including perivascular cells and smooth muscle cells (Fig. 1b). The CTSK+ cells covered a large portion of the tendon-derived mesenchymal stem cells (Fig. 1b). This remains the case in the late stages of injury (Fig. S1B). At the same time, pseudotime analysis of the CTSK+ cell population found that CTSK+ cells could differentiate into osteogenic and fibrogenic directions after injury (Fig. 1c). Pseudotime analysis of the PRRX1+ cell population found that, in addition to osteogenic and fibroblastic differentiation, PRRX1+ cells also differentiated toward myogenesis (Fig. 1c). Cell frequency analysis of several representative marker genes in the MSC population revealed that CTSK+ cells were more injury-responsive and widely distributed (Fig. 1d, Fig. S1E). Not only was the number of CTSK+ cells increased, but their overall expression was also greater than prrx1. In addition, we compared signature target genes in the osteogenic and fibrotic directions and found that the expression of osteogenic and fibrotic signature genes was significantly greater in CTSK+ cells than in CTSK- cells (Fig. 1d, Fig. S1D). We also found that the CTSK+ gene labeled more cells than TDPCs which just expressing tendon-specific genes (Fig. S1C, F).
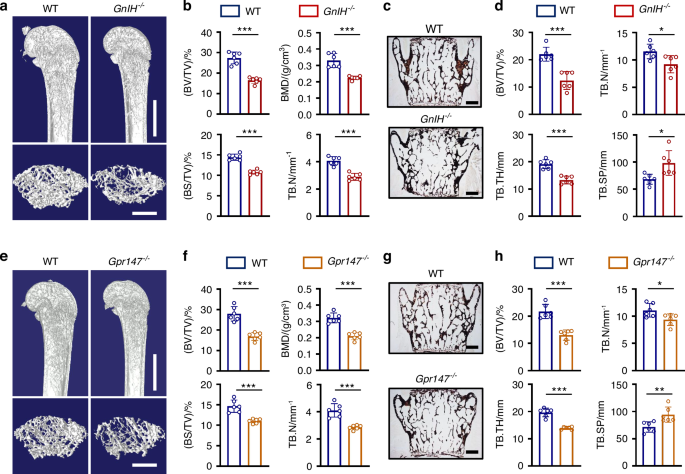
Fig. 1
CTSK lineage tendon-derived mesenchymal stem cells differentiate into osteogenic and fibrogenic directions at the Achilles tendon site after injury. a Single-cell sequencing data analysis of the Achilles tendon injury site, identifying 19 clusters, including mesenchymal stem cells based on Pdgfra and Prrx1 genes, where the cell population with higher expression of these two genes is defined as TDMSCs, macrophage based on Lyz2 and Cd68, etc. b Expression distribution of Ctsk+ and Prrx1+ cells in all the cells obtained by single-cell sequencing. c Trajectory analysis of gene expression changes across pseudotime in the Ctsk+ and Prrx1+cell cluster. d Cell frequency analysis of several representative marker genes in the TDMSC cell population, including Ctsk, Prrx1, Pdgfra, Nt5e, Nestin and LepR. e Representative images of immunofluorescence staining with anti-GFP and anti-OCN. Scale bars: 100μm. Cell nuclei were stained with DAPI. The Achilles tendon images at the HO site (top) were marked by white dashed lines at x20 magnification, and the images at x63 magnification (bottom) in the white dashed boxes were selected for statistical analysis of cell counting, showing the antibody staining channels separately. f Representative images of immunofluorescence staining with anti-GFP and anti-COL3 in the specimens of CTSK-traced mice 1 week after injury modeling. Scale bars: 100 μm. g Statistical comparison of the number of OCN+ cells, YFP+OCN+ and YFP+ COL3+ double-positive cells in the field of view in the images at x63 magnification. (n = 4/group, ****P < 0.000 1)
To further study the function and fate of CTSK lineage mesenchymal cells, we constructed transgenic CTSK-YFP lineage tracing mice with the HO mouse model (Fig. S1G, H). Micro-CT scanning and Masson’s trichrome staining revealed ectopic bone formation and bone marrow cavity in the Achilles tendon area (Fig. S1I, J). Further immunofluorescence staining revealed that many CTSK-YFP cells were costianed with osteocalcin (OCN, an osteoblast marker), indicating that CTSK+ cells or their subsequent differentiated cells formed osteogenic cells (Fig. 1e, g). To further study the differentiation of CTSK+ lineage cells into fibroblasts in situ, COL3+ cells, which represent newly formed fibers, were measured and showed costained with CTSK+ lineage cells in the injured Achilles tendon area (Fig. 1f, g), which revealed that CTSK+ cells can differentiate into COL3+ fibroblasts upon injury. These data from a lineage tracing mouse model and single-cell sequencing data suggest that CTSK- lineage TDMSCs are amajor source of oste oblastic and fibroblastic lineage cells after Achilles tendon injury.
The CTSK lineage tendon-derived mesenchymal stem cell population is sensitive to mechanical stimulationTo study the effects of mechanical stimulation on HO, we immobilized the distal limbs of the mice after Achilles tendon injury, so that the ankle joints of the mice were in plantar flexion (Fig. 2a). This fixed position placed the Achilles tendon and triceps surae in a relaxed state and under nonstretching stress. Moreover, the external immobilization device prevented pressure from being transmitted to the ankle joint when the mouse walked.19 We performed single-cell sequencing data analysis of the Achilles tendon area of mobile and immobile mice after injury modeling and found that with immobilizaztion, the CTSK lineage cell population was decreased compared with that in the mobile group (Fig. 2b, Fig. S2C). Additionally, CTSK+ TDMSCs were a more mechanosensitive and widely distributed cell subpopulation. A comparison of mobile and immobile samples also revealed that CTSK expression levels changed considerably with mechanical conditions (Fig. 2b, Fig. S2D). The GO analysis of the CTSK+ and CTSK- cell populations revealed alterations in several mechanical force-related pathways (Fig. 2c).
Fig. 2
CTSK lineage tendon-derived mesenchymal stem cell population is sensitive to mechanical stimulation. a Schematic diagram of immobilized mechanical intervention after Achilles tendon injury modeling by Burn/ATP. b Cell frequency analysis of several representative marker genes in the TDMSC cell population comparing Mobile and Immobile, including Ctsk, Prrx1, Pdgfra, Nt5e, Nestin and LepR. c GO analysis of Ctsk+ as well as Ctsk- cell populations revealed alterations in several mechanical force-related pathways. d, e Mice were subjected to mobile and immobile interventions after Burn/ATP modeling, and specimens were collected for MicroCT scanning, followed by decalcification and paraffin embedding of the specimens for masson immunohistochemical staining. Scale bars: 100 μm. f Atomic force microscopy scanning images of the ECM arrangement of the specimens of the mobile and immobile groups after injury. Scale bars: 1 μm. g Statistical analysis results of the bone volume of ectopic ossification shown by MicroCT scanning. Statistical analysis of the area of collagen fibers in the field of view of the masson immunohistochemical staining microscope images and Anisotropy analysis of AFM images of the mobile and immobile groups by independent samples t-test (n = 6/group, *P < 0.05, **P < 0.01, ****P < 0.000 1). h Representative images of immunofluorescence staining with anti-GFP and anti-OCN of the specimens of Ctsk-YFP lineage-traced mice divided into mobile and immobile groups after Burn/ATP modeling. Scale bars: 100 μm. i Representative images of immunofluorescence staining with anti-GFP and anti-COL3 of the specimens of Ctsk-YFP lineage-traced mice divided into mobile and immobile groups after Burn/ATP modeling. Scale bars: 100 μm. j Statistical comparison of the number of OCN+ and YFP+ cells in the field of view of the images, comparing the mobile and immobile groups by independent samples t-test (mobile group n = 6, immobile group n = 4, ***P < 0.001). k Statistical comparison of the number of COL3+ cells and COL3+YFP+ double-positive cells in the field of view of the images, comparing the mobile and immobile groups by independent samples t-test (mobile group n = 6, immobile group n = 4, ***P < 0.001)
To further verify the effect of mechanical stimulation on CTSK lineage cell function in vivo, we performed immobilized interventions on a CTSK-YFP lineage tracing mouse model after Burn/Achilles tendon puncture modeling. The volume of heterotopic ossification decreased after immobilization (Fig. 2d, g), and the number of collagen fibers at the connection between the Achilles tendon and calcaneus also decreased (Fig. 2e, g). Moreover, although the ultrastructure of the ECM appeared to be unaltered, the arrangement of the ECM became more disorganized after immobilization (Fig. 2f, g). We performed an in vitro tensile stress test on isolated TDMSCs and obtained similar results, with enhanced osteogenic and fibroblastic differentiation following stress. Immunofluorescence staining analysis revealed that the number of CTSK+ cell-derived osteoblast-like cells decreased after immobilization (Fig. 2h, j). To detect fibrogenic activity in the Achilles tendon area under different mechanical intervention conditions after injury, we performed immunofluorescence staining of newly formed fibrous COL3 and found that the number of cells expressing CSTK and COL3 decreased after immobilization (Fig. 2i, k).
Polycystin-1 mediated mechanical signals affect abnormal osteogenesis and ECM structures after Achilles tendon injuryTo further explore how CTSK+ cells sense mechanical signal transduction, we analyzed the expression levels of mechanical signal transduction related genes in CTSK+ cells via scRNA-seq analysis. Many types of mechanical signal transduction genes were expressed in CTSK+ cells. Among several common mechanical signal transduction genes, we found that the Pkd1 (encoding gene for polycystin-l (PC1)) and Wwtr1 (coding gene for TAZ) genes presented relatively high expression levels (Fig. 3a). In addtion, during immoblization, the expression of Pkd1 and Wwtr1 dramatically decreased (Fig. 3b). Moreover, Pkd1+ cells exhibited enhanced expression of osteogenic and fibrogenic genes. Additionally, the osteogenic or fibrogenic differentiation bias of Pkd1+ cells was observed to vary with different mechanical intervention conditions (Fig. S2E). Given that the PC1-TAZ axis has been shown to regulate bone formation and BMSC differentiation during skeletal modeling and remodeling,16 we hypothesized that CTSK+ cells mainly sense mechanical stimulation and affect HO through PC1.
Fig. 3
Polycystin-1 conducting mechanical signals affects abnormal osteogenesis and ECM structures after Achilles tendon injury. a Ctsk+ cells were isolated from all the cells obtained by single-cell sequencing, and the expression levels of some common mechanosensitive protein genes were analyzed, including Yap1 (YAP), Wwtr1 (TAZ), Ptk2 (FAK), Pkd1 (PC1), Piezo1 and Piezo2. b Ctsk+ TDMSCs were isolated from all the cells obtained by single-cell sequencing, and compared for the effects of mobile or immobile interventions after HO modeling on mechanoreception-related genes expressed in these cells, including Pkd1, Wwtr1, Yap1, Ptk2, Piezo1 and Piezo2. c Violin plot of Pkd1 gene expression level in Ctsk+ and Ctsk- cells. d Representative images of immunofluorescence staining with anti-GFP and anti-PC1 of the specimens of Ctsk-YFP lineage-traced mice divided into mobile and fixed groups after Burn/ATP modeling. Cell nuclei were stained with DAPI. The Achilles tendon images at the HO site (top) were marked by white dashed lines at x20 magnification, and the images at x63 magnification (bottom) in the white dashed boxes were selected for statistical analysis of cell counting, showing the antibody staining channels separately. e Schematic diagram of the experiment to induce local Pkd1 knockout by AAV-Cre injection. f Representative 3D reconstruction images of MicroCT scanning of the specimens of Pkd1fl/fl mice injected with AAV-Ctrl and AAV-Cre (2× 1010 plaque-forming units per mouse) after Burn/ATP modeling, and statistical analysis results of the bone volume of ectopic ossification shown by MicroCT scanning. g Atomic force microscopy scanning images and Anisotropy analysis of the ECM arrangement of the specimens of AAV-Ctrl and AAV-Cre mice after Burn/ATP modeling. Scale bars: 1 μm. h Representative images of immunofluorescence staining with anti-PC1 and anti-OCN of the HO site of the specimens of AAV-Ctrl and AAV-Cre mice after Burn/ATP modeling. Scale bars: 100 μm. i Statistical comparison of the number of PC1+ cells and OCN+ cells in the field of view of the images, comparing the Ctrl group and the AAV-Cre injection group by independent samples t-test (n = 6, ***P < 0.001, ****P < 0.000 1). j Representative images of immunofluorescence staining with anti-COL3 of the Achilles tendon site of the specimens of AAV-Ctrl and AAV-Cre mice after Burn/ATP modeling. Scale bars: 100 μm. k Statistical comparison of the number of COL3+ cells in the field of view of the images, comparing the Ctrl group and the AAV-Cre injection group by independent samples t-test (n = 6, ****P < 0.000 1)
To further verify the role of the PC1 protein in local mechanical signal transduction after Achilles tendon injury in vivo, we performed immobilization interventions on a CTSK lineage tracing mouse model. Immunofluorescence staining revealed that in the mobile group, most CTSK-lineage cells were costianed with PC1 in the cell membrane, which significantly decreased with immobilizaiton (Fig. 3d, Fig. S1I). To test the function of PC1 in HO, we injected the AAV-Cre virus into the Achilles tendon of Pkd1fl/fl mice after Burn/ATP Achilles tendon injury (Fig. 3e). After the virus was injected, Cre gene expression was induced in local cells of injured Achilles tendon, thereby achieving local inducible gene knockout in Pkd1fl/fl mice. Compared with that in the control group, the amount of ectopic ossification decreased with AAV-Cre treatment (Fig. 3f), and the structure of the ECM thus became disorganized(Fig. 3g). Immunofluorescence staining of tissue sections revealed that PC1 protein and OCN+ osteoblast expression was reduced after AAV-Cre was injected into Pkd1fl/fl mice (Fig. 3h, i). Compared with those in the control group, the number of COL3+ fibroblasts in the injured area was decreased in the AAV-Cre-injected group (Fig. 3j, k).
Conditional knockdown of Pkd1 in TDMSCs alters aberrant osteogenesis and the ECM in vivoThe use of the AAV-Cre virus for inducible gene knockout results in the knockout of Pkd1 in all local cells. To avoid any confounding effects, we created a CTSK+ cell conditional Pkd1 gene-specific knockout mouse model, Ctsk-Cre+/−-Pkd1fl/fl CKO mice, to confirm the role of the PC1 protein in CTSK+ cells in vivo. Pkd1fl/fl litters served as control (Fig. 4a). We established the same Burn/ATP injury model in WT and CKO mice. After the specific knockout of the Pkd1 gene in CTSK+ cells, HO formation was significantly reduced (Fig. 4b, i). The arrangement of the ECM was also disorganized (Fig. 4c, i). In addition, the number of osteoblast-like cells and newly formed fibroblast-like cells at the injury site was decreased in CKO mice (Fig. 4d, f, g).
Fig. 4
Conditional knockdown of Pkd1 in TDMSCs alters aberrant osteogenesis and ECM in vivo. a Schematic diagram of the conditional gene knockout mouse with Ctsk+ cell-specific knockout of Pkd1. b Representative 3D reconstruction images of MicroCT scanning of the specimens of CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. c Atomic force microscopy scanning images of the ECM arrangement of the specimens of CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. Scale bars: 1 μm. d Representative images of immunofluorescence staining with anti-PC1 and anti-OCN of the HO site of the specimens of CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. Cell nuclei were stained with DAPI. Scale bars: 100 μm. e, f Statistical comparison of the number of PC1+ cells and OCN+ cells in the field of view of the images, comparing CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice by independent samples t-test (n = 6, **P < 0.01). g Representative images of immunofluorescence staining with anti-COL3 of the Achilles tendon site of the specimens of CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. Scale bars: 100 μm. h Statistical comparison of the number of COL3+ cells in the field of view of the images, comparing CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice by independent samples t-test (n = 6, ***P < 0.001). i Statistical analysis results of the bone volume of ectopic ossification shown by MicroCT scanning, analyzing total HO Bone Volume, and Anisotropy analysis of AFM images. j Schematic diagram of the conditional gene knockout mouse with Prx1+ cell-specific knockout of Pkd1. k Representative 3D reconstruction images of MicroCT scanning of the specimens of Prx1Cre−/−Pkd1fl/fl control mice and Prx1Cre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. l Atomic force microscopy scanning images of the ECM arrangement of the specimens of Prx1Cre−/−Pkd1fl/fl control mice and Prx1Cre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. Scale bars: 1 μm. m Representative images of immunofluorescence staining with anti-PC1 and anti-OCN of the HO site of the specimens of Prx1Cre−/−Pkd1fl/fl control mice and Prx1Cre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. Scale bars: 100 μm. n, o Statistical comparison of the number of PC1+ cells and OCN+ cells in the field of view of the images, comparing Prx1Cre−/−Pkd1fl/fl control mice and Prx1Cre+/−Pkd1fl/fl knockout mice by independent samples t-test (n = 4, *P < 0.05). p Representative images of immunofluorescence staining with anti-COL3 of the Achilles tendon site of the specimens of Prx1Cre−/−Pkd1fl/fl control mice and Prx1Cre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. Scale bars: 100 μm. q Statistical comparison of the number of COL3+ cells in the field of view of the images, comparing Prx1Cre−/−Pkd1fl/fl control mice and Prx1Cre+/−Pkd1fl/fl knockout mice by independent samples t-test (n = 4, **P < 0.01). r Statistical analysis results of the bone volume of ectopic ossification shown by MicroCT scanning, analyzing total HO Bone Volume, and Anisotropy analysis of AFM images
We next generated Prrx1-Cre+/−-Pkd1fl/fl conditional gene knockout mice (Fig. 4j), to confirm the effect of PC1 on TDMSC mechanical-signal transduction in vivo. The Prrx1 gene encodes a paired-related homeobox 1 (PRRX1) protein, which is a transcription factor expressed in multiple tissues, especially in neural crest and limb buds, and is generally considered a classical mesenchymal stem cell marker.20 After the same Burn/ATP injury model was established in Prrx1-Cre+/−-Pkd1fl/fl and Prrx1-Cre−/−-Pkd1fl/fl mice, micro-CT scanning was performed to measure the amount of ectopic ossification. We found that the HO volume in the CKO group was significantly lower than that in the WT group (Fig. 4k, r). Pkd1 gene knockout in PRRX1+ cells impaired mesenchymal cell osteogenic and fibrogenic differentiation (Fig. 4m–q), and resulted in decreased anisotropy and a poor ultrastructure of the ECM (Fig. 4l, r). These data demonstrate that PC1 regulates aberrant TDMSC osteogenic and fibrogenic fates and ECM organization.
Polycystin 1 transmits mechanical signals through downstream TAZPrevious studies have shown that PC-1 transduces mechanical signals through the downstream transcriptional coactivator TAZ.15,16 Earlier studies revealed that TAZ plays an important role in promoting the differentiation of MSCs into osteoblasts and inhibiting their differentiation into adipocytes. TAZ can bind to Runx2 to form a TAZ-Runx2 complex, which in turn transcriptionally activates osteogenic genes, such as Osterix and Osteocalcin.21 Single-cell analysis data also revealed elevated expression of osteogenic and fibrogenic genes in Taz+ cells. Moreover, the propensity of Taz+ cells to differentiate into either osteoblasts or fibroblasts was found to be modulated by varying mechanical stimuli. We then tested whether the PC1-TAZ axis also influences cell functions in a HO mouse model. We isolated TDMSCs from murine Achilles tendons following previous methods,22,23,24 and verified the function of PC1 in vitro (Fig. 5a). The osteogenic activity of tendon mesenchymal stem cells, as measured by ALP staining, decreased after Pkd1 was silenced with siRNA(Fig. 5b). TAZ nuclear translocation, which mediates downstream signaling, decreased after Pkd1 was knocked down in contrast to the control group (Fig. 5c). These findings indicate that PC1 may regulate mechanical signal transduction and osteogenic potential in tendon-derived mesenchymal stem cells through TAZ nuclear translocation.
Fig. 5
Polycystin 1 transmits mechanical signals through downstream TAZ in Tendon-derived MSCs. a Schematic diagram of the extraction of MSCs at the Achilles tendon site. b Representative images of ALP staining of Achilles tendon mesenchymal cells after PKD1 siRNA interference, compared with NC siRNA. c Separation of cytoplasmic and nuclear proteins of TDMSCs after PKD1 siRNA interference to detect the nuclear translocation of TAZ, compared with Si NC group. Biological repetitions = 3. d Representative images of immunofluorescence staining with anti-GFP and anti-TAZ of the specimens of Ctsk-YFP lineage-traced mice divided into mobile and immobile groups after Burn/ATP modeling. Cell nuclei were stained with DAPI. Scale bars: 100 μm. e Statistical comparison of the number of TAZ+ cells and TAZ nuclear translocation in the field of view of the images, comparing the mobile and fixed groups by independent samples t-test (mobile group n = 6, fixed group n = 4, ***P < 0.001). f Representative images of immunofluorescence staining with anti-PC1 and anti-TAZ of the HO site of the specimens of CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice after Burn/ATP modeling. Scale bars: 100 μm. g Statistical comparison of the number of TAZ nuclear translocation cells in the field of view of the images, comparing CtskCre−/−Pkd1fl/fl control mice and CtskCre+/−Pkd1fl/fl knockout mice by independent samples t-test (n = 6, **P < 0.01)
To further investigate the role of the PC1-TAZ mechanical signal transduction pathway in the complex environment of tendon injury in vivo, we generated Burn/ATP injury models in a CTSK-YFP lineage tracing mouse model and divided them into mobile and immobile groups. The results of immunofluorescence staining indicated that CTSK-lineage cells colocalized with TAZ at the injury site. Interestingly, the fluorescence signal of TAZ was more concentrated in the cytoplasm in the immobilized group than in the mobilized control group (Fig. 5d, e), indicating that immobilization suppresses TAZ nuclear translocation. More importantly, the nuclear localization of TAZ was also lower in Ctsk-Cre+/−-Pkd1fl/fl CKO mice than in Pkd1fl/fl littermate controls (Fig. 5f, g).
To further confirm the role of the PC1-TAZ pathway in HO and the ECM after tendon injury, DAPT, a γ-secretase inhibitor that inhibits the cleavage of the PC1-CTT terminal and thus inactivates the PC1-TAZ signaling pathway, was used in subsequent experiments.16,25 DAPT treatment reduced the osteogenic activity of tendon-derived mesenchymal stem cells in vitro, as evidenced by decreased ALP activity (Fig. 6a). Additionally, DAPT may also simultaneously affect the Notch pathway. To investigate through which pathway DAPT mainly exerts its influence, we further compared the inhibitory effect of DAPT on TDMSCs with that of IMR-1, a specific inhibitor of the Notch pathway. After IMR-1 inhibited the Notch pathway, DAPT had a further inhibitory effect, suggesting that DAPT v has targets other than the Notch pathway. Moreover, after we overexpressed membrane-bound PC1-CTT, its overexpression was inhibited by DAPT. This finding also indicates, to some extent, that DAPT does play a role by affecting PC1-CTT. We then tested the effect of DAPT in vivo by local subcutaneous injection of DAPT around the tendon injury site in Burn/ATP HO model mice treated with 10 mg/kg DAPT twice a week (Fig. 6b). Compared with vehicle treatment, DAPT treatment dramatically suppressed HO formation (Fig. 6c, d), and reduced the number of osteoblast-like cells and fibroblast-like cells in injured tendons (Fig. 6e, f). As expected, the localization of TAZ to the nucleus was inhibited by DAPT administration (Fig. 6g). These data suggest that PC1-CTT cleavage-induced TAZ nuclear translocation promotes the osteogenic and fibrogenic differentiation of MSCs and HO progression.
Fig. 6
Injection of PC1-CTT inhibitor DAPT attenuates osteogenic and fibrogenic activity after injury. a Representative images of ALP staining of Achilles tendon mesenchymal stem cells after DAPT intervention, compared with Vehicle. b Schematic diagram of the mouse injected with DAPT after injury modeling. c Representative 3D reconstruction images of MicroCT scanning of the specimens of WT mice injected with Vehicle or DAPT (10 mg/kg) after Burn/ATP modeling. d Statistical analysis results of the bone volume of ectopic ossification shown by MicroCT scanning. e Representative images of immunofluorescence staining with anti-OCN of the HO site of the specimens of Ctrl mice injected with Vehicle and mice injected with DAPT after Burn/ATP modeling. Cell nuclei were stained with DAPI. Scale bars: 100 μm. f Representative images of immunofluorescence staining with anti-COL3 of the Achilles tendon site of the specimens of Ctrl mice injected with Vehicle and mice injected with DAPT after Burn/ATP modeling. Scale bars: 100 μm. g Representative images of immunofluorescence staining with anti-TAZ of the Achilles tendon site of the specimens of Ctrl mice injected with Vehicle and mice injected with DAPT after Burn/ATP modeling. Scale bars: 100 μm. h Statistical comparison of the number of OCN+, Nuclear TAZ+, and COL3+ cells in the field of view of the images, comparing Ctrl mice injected with Vehicle and mice injected with DAPT by independent samples t-test (n = 6, **P < 0.01, ***P < 0.001, ****P < 0.000 1)
Conditional knockdown of Taz in TDMSCs alters aberrant osteogenesis and the ECM in vivoTo further verify the role of TAZ in the mechanical signal transduction pathway in CTSK+ cells after tendon injury, we crossed Ctsk-Cre+/− mice with Tazfl/fl mice to generate CTSK+ cell conditional Taz gene specific knockout mice, followed by Achilles tendon injury Tazfl/fl litters served as controls (Fig. 7a). First, the HO phenotype was analyzed. Compared with Tazfl/fl mice, HO formation at both the proximal and distal sites was significantly lower in Ctsk-Cre+/−-Tazfl/fl mice (Fig. 7b, c). Immunofluorescence staining analysis revealed that both osteoblast-like cells and fibroblast-likes cells were diminished in Ctsk-Cre+/−-Tazfl/fl mice (Fig. 7d, e). We verified the depletion of Taz in CTSK+ cells by detecting the TAZ fluorescence signal and detected a significantly lower TAZ signal in situ (Fig. 7f). Moreover, ECM alignment became disorganized with TAZ depletion (Fig. 7h). These data indicate that TAZ in CTSK+ cells is required for HO formation and ECM organization and that blocking TAZ is sufficient to prevent or alleviate HO.
Fig. 7
Conditional knockdown of Taz in CTSK+ TDMSCs alters aberrant osteogenesis and ECM in vivo. a Schematic diagram of the conditional gene knockout mouse with Ctsk+ cell-specific knockout of Taz. b Representative 3D reconstruction images of MicroCT scanning of the specimens of CtskCre−/−Tazfl/fl control mice and CtskCre+/−Tazfl/fl knockout mice after Burn/ATP modeling. c Statistical analysis results of the bone volume of ectopic ossification shown by MicroCT scanning, analyzing total HO Bone Volume. d Representative images of immunofluorescence staining with anti-OCN of the HO site of the specimens of CtskCre−/−Tazfl/fl control mice and CtskCre+/−Tazfl/fl knockout mice after Burn/ATP modeling. Cell nuclei were stained with DAPI. Scale bars: 100 μm. e Representative images of immunofluorescence staining with anti-COL3 of the Achilles tendon site of the specimens of CtskCre−/−Tazfl/fl control mice and CtskCre+/−Tazfl/fl knockout mice after Burn/ATP modeling. f Representative images of immunofluorescence staining with anti-TAZ of the Achilles tendon site of the specimens of CtskCre−/−Tazfl/fl control mice and CtskCre+/−Tazfl/fl knockout mice after Burn/ATP modeling. g Statistical comparison of the number of OCN+, TAZ+, and COL3+ cells in the field of view of the images, comparing CtskCre−/−Tazfl/fl control mice and CtskCre+/−Tazfl/fl knockout mice by independent samples t-test (n = 4, *P < 0.05, **P < 0.01, ***P < 0.001). h Atomic force microscopy scanning images of the ECM arrangement of the specimens of CtskCre−/−Tazfl/fl control mice and CtskCre+/−Tazfl/fl knockout mice after Burn/ATP modeling. Scale bars: 1 μm
These results indicate that mechanical force stimulates osteogenesis and fibrogenesis, thereby influencing heterotopic ossification and matrix organization, via PC1-CTT and TAZ nuclear translocation in tendon-derived mesenchymal stem cells (Fig. 8).
Fig. 8
Schematic representation of TDMSCs sensing mechanical signals through the PC1-TAZ axis thus osteogenic and fibroblastic differentiation affecting ECM and HO. Mechanical force promotes osteogenesis and fibrogenesis through PC1-CTT and TAZ nuclear translocation in tendon-derived mesenchymal stem cells
Comments (0)