
Remember me


Various studies have shown that overexpression of the ER-associated USP19 ubiquitin peptidase promotes unconventional secretion of misfolded proteins such as α-synuclein, into the extracellular medium via the MAPS pathway [26, 28, 30]. Conversely, no secretion of misfolded proteins was observed when a cytosolic form of USP19, deleted for its transmembrane domain necessary for ER-anchoring (USP19-ΔTM; see Fig. 1a), was overexpressed. These data indicate the crucial role of its association to the ER membrane in the secretion process [26, 28]. Interestingly, Xu et al., (2018) also observed that co-expression of USP19 with wild type TDP-43, barely increased the amount of TDP-43 in the extracellular medium [28]. These data prompted us to investigate the possibility that ER-USP19 could specifically promote the secretion of misfolded TDP-43 mutants prone to aggregation. In this context, various combinations of WT and mutant TDP-43 and USP19 were overexpressed in HEK293T cells. We first overexpressed untagged-TDP-43-WT and the familial TDP-43-K263E ALS variant [36] (Fig. 1b). Immunofluorescence experiments using anti-TDP-43 antibodies confirmed that wild type TDP-43 was predominantly expressed in the nucleus with a slight cytoplasmic distribution, whereas the K263E variant was distributed both in the nucleus and in the cytoplasm where large inclusion bodies could be observed (see white arrows in Fig. 1b). The presence of TDP-43 aggregates was confirmed by Western blotting after sarkosyl fractionation: a significant amount of TDP-43-K263E mutant was present into the insoluble fractions (Sark-ins) whereas only a faint signal was observed with TDP-43-WT (compare lanes 5 and 6 in Fig. 1c).
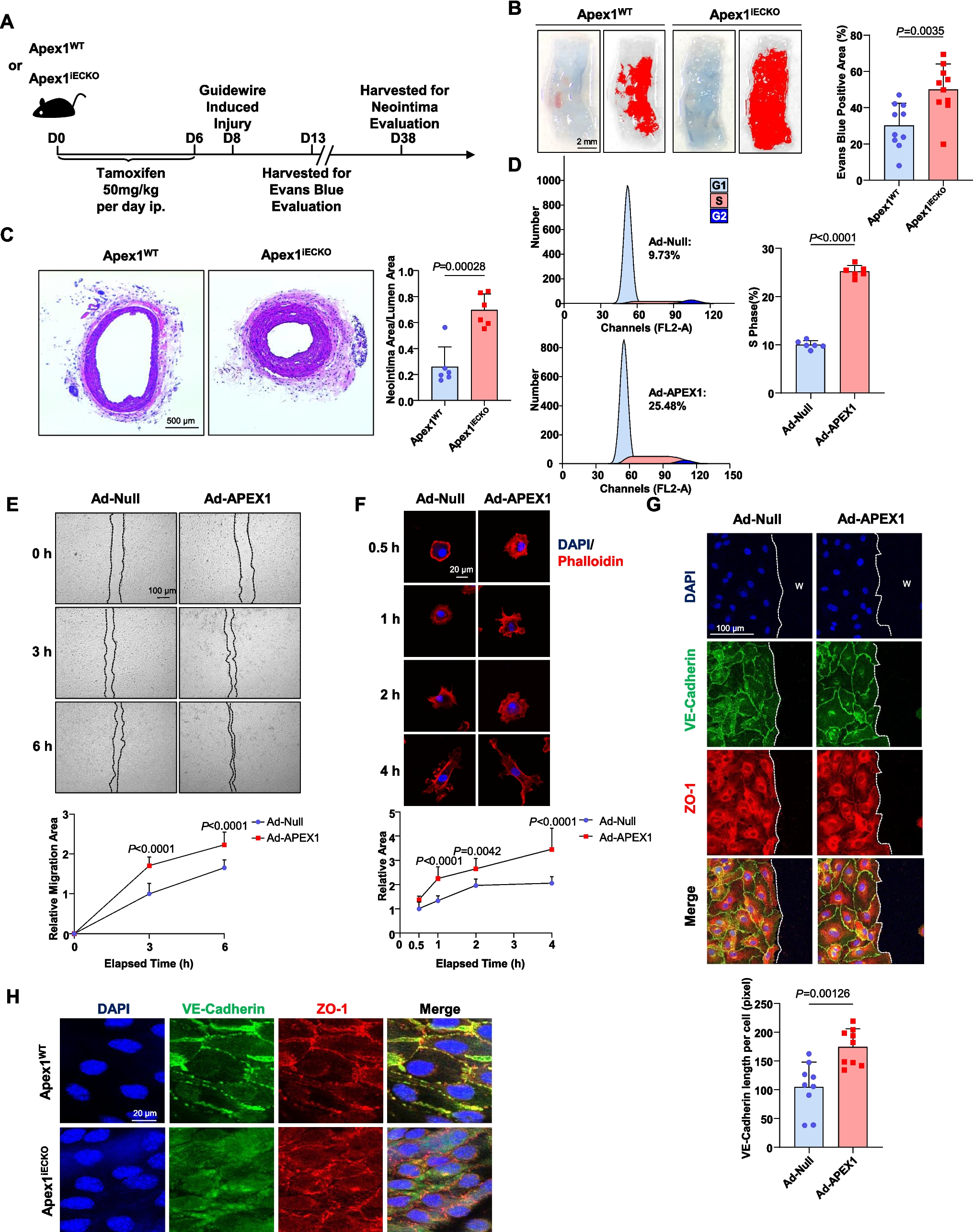
Fig. 1
The ER-anchored USP19 promotes the secretion of misfolded TDP-43 in the HEK293T cellular model. a Schematic representation of Flag-USP19 constructs used in this study. The C506 residue represents an essential amino acid residue necessary for the ubiquitin peptidase catalytic activity. CS 1&2 CHORD-containing proteins and STG1, UBL ubiquitin-like, USP ubiquitin-specific peptidase, TM transmembrane domain, ZnF zinc finger, ΔTM deleted for the transmembrane domain. The pink star represents the amino terminal Flag tag. b Schematic representation of the full-length TDP-43 (upper panel). The K263E variant is highlighted in blue. LCD Low Complexity Domain, NLS nuclear localization sequences, NTD N-terminal domain, RRM1&2 RNA recognition motif 1&2. HEK293T cells were transfected with TDP-43-WT or TDP-43-K263E encoding constructs and were visualized by immunofluorescence using anti-TDP-43 antibody. Blue signal corresponds to DAPI for nuclei and green signal to TDP-43. Scale bar is 10 µm. c Immunoblotting of sarkosyl soluble supernatant (Sark-sol) and sarkosyl insoluble pellet (Sark-ins) fractions isolated from control HEK293T cells (lanes 1 and 4), or cells expressing TDP-43-WT (lanes 2 and 5) or TDP-43-K263E (lanes 3 and 6) using antibodies directed against TDP-43 and GAPDH as loading control. d Evaluation of misfolded TDP-43 secretion by filter trap assay (FTA). Presence of misfolded TDP-43 in conditioned media from HEK293T cells overexpressing TDP-43-WT and TDP-43-K263E and the different Flag-USP19 or the empty vector (negative control) was monitored by FTA. Upper panel (secretion/conditioned media): nitrocellulose membranes were probed with an antibody directed against TDP-43. Lower panel (cell expression): Western blotting of cell lysates from co-expressing cells using anti-Flag (for USP19), -TDP-43 and -GAPDH antibodies for loading control. e Quantification of secreted TDP-43 upon USP19 expression. Data represent mean ± SEM, n = 5 experiments. Significance was assessed by a Mann–Whitney U test (**p < 0.001). f USP19-WT or USP19-ΔTM and TDP-43-K263E overexpression does not affect plasma membrane permeability and cell viability. HEK293T cells transfected with the indicated plasmids were stained with trypan blue and counted. Data represent mean ± SEM, n = 3 experiments
As expected, Flag-USP19-WT strongly accumulated in the ER as evidenced by colocalization with the KDEL ER-marker. In contrast, the USP19-ΔTM mutant (Fig. 1a), lacking the transmembrane domain necessary for ER-anchoring did not colocalize with KDEL labeling (see Fig. S1a, b). Neither USP19-WT nor USP19-ΔTM colocalized with the Golgi apparatus marker GM130 (Fig. S1c, d).
Based on these data, we next investigated the impact of flagged USP19-WT and ΔTM on TDP-43-WT secretion in co-expressing cells. To evaluate TDP-43 secretion, conditioned media were analyzed by filter trap assay (FTA) in presence of 1% SDS detergent to specifically detect insoluble TDP-43 aggregates. The results presented in Fig. 1d, e showed that USP19-WT only slightly promoted the secretion of TDP-43-WT aggregates compared to the USP19-ΔTM, corroborating previously published data [28]. Conversely, when co-expressed with the pathological TDP-43-K263E (Fig. 1b), USP19-WT, but not the USP19-ΔTM, significantly increased the secretion of aggregated TDP-43 (Fig. 1d, e). Similar results were obtained when HA-Tagged TDP-43-K263E was co-expressed with USP19-WT in the presence (Fig. S2a-c) or absence of endogenous USP19 expression (Fig. S2d, e).
Interestingly, Xu et al., (2018) observed that the C-terminal region of USP19 encompassing amino acid residues 494–1318 (USP19494−1318; see Fig. 1a), which include the Ubiquitin peptidase activity and the ER-anchoring transmembrane domain, was sufficient to enhance the secretion of the misfolded proteins including α-synuclein [28]. In contrast, overexpression of the N-terminal part of USP19 (USP191−493; Fig. 1a) did not induce secretion [26]. These data prompted us to evaluate the impact of both truncated forms of USP19 on TDP-43-WT and TDP-43-K263E secretion. The data presented in Fig. 1d,e, confirmed that USP19494−1318 promoted the secretion of misfolded TDP-43-WT and K263E, while the N-terminal USP191−493 did not enhance the release. This indicates that the 494–1318 domain of USP19 contains the essential functional properties for misfolded TDP-43 secretion.
To ensure that the observed secretion of misfolded TDP-43 was not simply the reflect of cell death potentially induced by the combined expression of USP19-WT and TDP-43-K263E, we assessed cell viability using the trypan blue exclusion method. The data presented in Fig. 1f and S2c revealed comparable levels of viable cells across the conditions. These findings, indicate that the secretion of misfolded TDP-43 was a specific cellular process rather than a byproduct of increased cell mortality.
Increased secretion of misfolded TDP-43 in the presence of USP19-WT or the C-terminus USP19494−1318 was confirmed with the cytosolic GFP-tagged TDP-43-ΔNLSΔ187-192 [35] mutant (Fig. S3a-c). Interestingly, when the same experiments were conducted with the C-terminus prion-like domain (GFP-TDP-43-CTF219−414) prone to high aggregation [35] (see Fig. S3a), neither USP19-WT nor its mutants stimulated TDP-43 secretion (Fig. S3d).
To validate our findings in a neuron-like model, we investigated the impact of USP19-WT on TDP-43-K263E secretion in the human neuroblastoma SH-SY5Y cell line. The data presented in Fig. S4a, b confirmed that overexpression of USP19-WT promotes the secretion of the misfolded TDP-43-K263E in this neuronal cellular model.
Overall, the data correlate well with those previously published on other prion-like proteins and indicate that the ER-anchored USP19 can promote the secretion of ALS TDP-43 misfolded mutants in different cellular models.
USP19 ubiquitin peptidase activity is essential for TDP-43 secretionUbiquitinated and hyperphosphorylated TDP-43 was identified as a primary constituent of the mislocalized and insoluble cytoplasmic inclusions in the ALS and ALS-FTLD affected brains [3, 4]. Recently, a growing body of evidence indicated that ubiquitinating and deubiquitinating pathways are critically engaged in the fate decision of aberrant or pathological TDP-43 proteins [41]. The data presented above revealed that USP19-WT promotes the secretion of misfolded TDP-43 but the mechanisms by which USP19-WT induces this secretion are still unresolved. In this context, we first evaluated the involvement of the USP19 ubiquitin peptidase activity on the misfolded TDP-43-K263E secretion. For this purpose, the ubiquitin peptidase catalytically inactive USP19-C506S mutant [28] was co-expressed with the TDP-43-K263E mutant in HEK293T cells (Fig. 2a). Western blotting on whole cell extracts showed that expression of the USP19-C506S mutant increased the levels of ubiquitinated proteins compared to USP19-WT and USP19-ΔTM (compare lanes 1&2 with lane 3 in Fig. 2b). Analysis of the culture medium by FTA revealed that the increase in protein ubiquitination was associated with a loss of TDP-43-K263E secretion, suggesting that beyond its association with the ER membrane, USP19 ubiquitin peptidase activity is crucial for the secretion of misfolded TDP-43 (Fig. 2c, d).
Fig. 2
The ubiquitin peptidase activity is essential for TDP-43-K263E secretion in USP19 context. a Schematic representation of Flag-USP19 constructs. The USP19-C506S corresponds to an ER-anchored but ubiquitin peptidase catalytically inactive mutant. b Ubiquitination level in cell lysates of co-expressing cells. Immunoblotting of cell lysates from TDP-43-K263E and the indicated Flag-USP19 co-expressing cell using antibodies directed against Ubiquitin or GAPDH as loading control. c Evaluation of misfolded TDP-43 secretion by FTA. Presence of misfolded TDP-43 in conditioned media from HEK293T cells overexpressing the TDP-43-K263E, the WT, the ΔTM or the C506S Flag-USP19, was monitored by FTA. Upper panel (secretion/conditioned media): nitrocellulose membranes were probed with an antibody directed against TDP-43. Lower panel (cell expression): Western blotting of cell lysates from co-expressing cells using anti-Flag (for USP19), -TDP-43 and -GAPDH antibodies for loading control. d Quantification of secreted TDP-43-K263E upon USP19s expression. Data represent mean ± SEM, n = 5 experiments. Significance was assessed by a Mann–Whitney U test (**p < 0.001). e TDP-43-K263E deubiquitination is promoted by USP19-WT. Lysates from HEK293T cells co-expressing Myc-TDP-43-K263E and HA-Ubiquitin-WT with Flag-USP19-C506S (ubiquitin peptidase deficient mutant; lane 1) or Flag-USP19-WT (lane 2) were immunoprecipitated with anti-Myc and immunoblotted with anti-HA and anti-Myc. Whole cell lysates (WCL; 5 μg/input) from co-expressing cells were immunoblotted using anti-Myc, anti-Flag and anti-GAPDH for loading control. Asterisks correspond to heavy and light chains of immunoglobulins used during the immunoprecipitation
To determine if misfolded TDP-43-K263E is ubiquitinated in our cellular system and requires to be deubiquitinated by USP19-WT for its secretion, co-immunoprecipitation experiments were conducted on lysates from HEK239T cells co-expressing a Myc-tagged-TDP-43-K263E, an HA-tagged-Ubiquitin-WT, and either the Flag-USP19-WT (promoting TDP-43 secretion) or the deubiquitinase-deficient Flag-USP19-C506S (no TDP-43 secretion context). Myc-TDP-43-K263E was immunoprecipitated using an anti-Myc antibody, and the immunoprecipitate was analyzed by Western blotting using an anti-HA antibody to detect the linked HA-Ubiquitin-WT. The data presented in Fig. 2e confirmed the ubiquitination of Myc-TDP-43-K263E when the Flag-USP19-C06S mutant was expressed (Lane 1), compared to lower ubiquitination levels when Flag-USP19-WT was expressed (Lane 2).
Overall, these findings support the conclusion that TDP-43-K263E deubiquitination and secretion is promoted by the USP19-WT expression.
USP19 overexpression promotes the secretion of soluble and free TDP-43 aggregatesMany studies have suggested that misfolded TDP-43 can be transmitted from cell to cell in a prion-like manner and contributes to the spreading of the pathology in different region of the central nervous system [13, 15, 16, 18,19,20]. Understanding the molecular and cellular mechanisms involved in the secretion as well as the transmission of pathological TDP-43 from cell to cell is of utmost importance to identify potential therapeutical targets. In this context, we tested whether USP19 overexpression can contribute to the cell–cell transmission of misfolded-TDP-43. For this purpose, conditioned medium from donor HEK293T cells co-expressing USP19-WT and HA-TDP-43-K263E were added to naïve HEK293T cells as previously depicted [42]. The data presented in Fig. 3a-c revealed that treatment of naïve HEK293T recipient cells with conditioned media from HEK293T donor cells co-expressing USP19-WT and HA-TDP-43-K263E showed a strong HA-TDP-43-K263E signal in recipient cells whereas only a very faint signal HA-TDP-43 was detected when the USP19–ΔTM mutant was expressed instead of USP19-WT in donor cells (Fig. 3b, c). These data suggest that USP19-WT overexpression may play a role in the intercellular transmission of secreted misfolded TDP-43 (Fig. 3a-c).
Fig. 3
The ER-anchored USP19 promotes the secretion of free TDP-43-K263E fibrils in the conditioned medium. a Transmission of misfolded HA-TDP-43-K263E to naïve HEK293T cells. Naive HEK293T cells were cocultured with conditioned media from HEK293T co-expressing cells Flag-USP19-WT or − ΔTM with HA-TDP-43-K263E during 72 h. b After 3 days, cells were washed and analyzed by Western blotting using anti-HA and anti-GAPDH as loading control. c Quantification of n = 5 independent experiments. Significance was assessed by a Mann–Whitney U test (**p < 0.001). d Aggregated TDP-43 levels in conditioned media from HEK293T cells overexpressing TDP-43-K263E and the Flag-USP19 WT and ΔTM were precleared to eliminate cellular debris and ultracentrifuged at 120,000 g pellet (p120K pellet) and analyzed by immunoblotting (lanes 3 and 4) using antibody directed against TDP-43. Cellular lysates (lanes 1 and 2) were analyzed by Western blotting and probed by antibodies directed against TDP-43, Flag and GAPDH (loading control). e Sucrose equilibrium density gradient fractionation of conditioned medium. The p120K pellet was fractionated through a linear sucrose equilibrium density gradient 9–60% and fractions (× 15), recovered from the top of the gradient, were analyzed by Western blotting using anti-TDP-43 or anti-CD81 and anti-CD63 antibodies for extracellular vesicles markers. Fraction density values (g/cm3) are depicted at the bottom panel. Whole cell lysate (WCL) of co-expressing cells was immunoblotted in parallel using the anti-TDP-43, anti-CD63 and anti-CD81. The bottom panel corresponds to the TCE staining of the SDS-PAGE gel. f Immunogold electron microscopy (IEM) of TDP-43-K263E positive fractions. Positive fractions (10–12) containing the TDP-43-K263E were pooled and analyzed by IEM using anti-TDP-43 labelled with a secondary antibody coupled with 10 nm gold particle. Amorphous and fibrillar structures were labelled (red arrows). Scale bars are 20 and 50 nm
Today, it is currently accepted that pathological TDP-43 can be released into the extracellular environment through various pathways, including extracellular vesicles (exosomes and/or microvesicles) [13, 19, 22,23,24] and/or as free aggregates [25]. To characterize in which form TDP-43-K263E was secreted upon USP19-WT overexpression, conditioned media from cells co-expressing USP19-WT and TDP-43-K263E were collected, centrifuged to remove cellular debris and ultracentrifuged at 120,000xg to pellet vesicles and protein aggregates (referred to as p120K). Secreted misfolded TDP-43 was detected in the p120K pellet, but not in the USP19–ΔTM context (Fig. 3d), confirming the previous FTA experimental results.
The p120K pellet was then fractionated through a linear 9–60% sucrose density gradients. Fifteen fractions were collected from the top, and analyzed by immunoblotting using antibodies against TDP-43 and the CD63 and CD81 extracellular vesicles (EVs) markers (Fig. 3e). While a faint TDP-43 signal was observed in the CD63 and CD81 positive fractions 6 and 7, the majority of TDP-43 was detected in a second peak, in fractions of higher densities (fractions 10 to 12; Fig. 3e). No TDP-43 signal was detected in any gradient fractions of the USP19–ΔTM context, although CD63 and CD81 EV markers were well detected (Fig. S5).
Immunogold electron microscopy (IEM) performed on TDP-43 positive fractions 10–12 revealed small fibrillar aggregated structures labeled with anti-TDP-43 and a secondary antibody coupled with 10 nm gold particles (indicated by red arrowheads in Fig. 3f). Similar observations were made when the GFP-TDP-43-ΔNLSΔ187-192 mutant was co-expressed instead of TDP-43-K263E with USP19-WT (Fig. S3e, f).
Given that MAPS pathway has been shown to enhance the secretion of soluble misfolded proteins into the extracellular space [26, 28], we next investigated if soluble TDP-43 could be detected in the conditioned medium after ultracentrifugation. To address this point, similar experiments were carried out as depicted above using HEK293T cells co-expressing Flag-USP19-WT and HA-TDP-43-K263E. The conditioned medium was collected and divided, with one third set aside as non-ultracentrifuged and the remainder subjected to ultracentrifugation at 120,000xg as previously depicted. Analysis of the p120K pellets confirmed the presence of TDP-43 aggregates when USP19-WT was expressed but not in presence of USP19-ΔTM (Fig. S6a). We then compared the ultracentrifuged supernatant with the non-ultracentrifuged sample using dot-blot assay without SDS detergent treatment to detect if soluble TDP-43 was secreted. The data presented in Fig. S6b, c revealed the presence of soluble TDP-43 in the resulting ultracentrifuged supernatant albeit at lower levels compared to non-ultracentrifuged condition. As anticipated, little or no TDP-43 signal was observed when USP19-ΔTM was expressed.
In conclusion, these findings indicate that USP19-WT expression promotes the secretion of both soluble and aggregated forms of misfolded TDP-43.
TDP-43 is associated with USP19 and the ER compartments and is engulfed in different intracellular compartments.To further characterize the cellular mechanisms involved in USP19-dependant trafficking and secretion of TDP-43-K263E, transmission electron microscopy (TEM) experiments were conducted. HEK293T cells co-transfected with both TDP-43-K263E and USP19-WT were compared to cells expressing TDP-43-K263E alone, TDP-43-K263E with USP19-ΔTM, or untransfected cells as negative control. Notably, cells co-expressing TDP-43-K263E and USP19-WT exhibited a significant accumulation of dilated ER (indicated by black arrows in panels i and v-ix in Fig. 4a). This observation correlated with the strong KDEL signal observed in Fig. S1a. No such ER accumulation was observed in the other conditions.
Fig. 4
Transmission electron microscopy (TEM) and RNA sequencing analyses point out ER alterations in TDP-43-K263E and USP19-WT co-expressing cells. a Ultrastructural analyses. TEM of TDP-43-K263E + UPS19-WT (panel i); TDP-43-K263E (panel ii); TDP-43-K263E + USP19-ΔTM (panel iii) and untransfected cells (UT; panel iv as negative control). Black arrows indicate dilated endoplasmic reticulum (ER) accumulation (panels i and v; v-right corresponds to a higher magnification of panel v). White asterisk indicates cytoplasmic aggregates (panel ii). White arrows indicate compact electron dense structures (CEDS; panel iii). Red arrows indicate mitochondria in close contacts with ER (panels v-right to vii). Yellow arrowheads indicate autophagic/endosomal compartments containing ER membrane and dense amorphous structures. (panels v-right, x and xi). Scale bars are 2 and 5 μm. Dotted squares correspond to higher magnification b Volcano plot of differentially expressed genes between TDP-43-K263E/USP19-WT versus TDP-43-K263E/USP19-ΔTM co-expressing cells. Red dots represents upregulated genes (P < 0.05 and Log2FC > 1), blue dots represents downregulated genes (P < 0.05 and Log2FC < -1), and grey dots represent genes that were not differentially expressed. c Bubble plot of the GO Biological process pathway enrichment analysis. The horizontal axis represents the enrichment score (-log10(p-value)), while the vertical axis represents the enriched pathway name. The color scale indicates different thresholds of the p-value, and size of the bubble indicates the number of genes corresponding to each pathway
RNA sequencing analyses performed on HEK293T cells co-expressing USP19-WT and TDP-43-K263E versus USP19-ΔTM and TDP-43-K263E revealed upregulated genes in USP19-WT co-expressing cells primarily associated with biological process linked to the ER including responses to topologically incorrect proteins, unfolded protein response, and, endoplasmic reticulum stress (Fig. 4b, c).
TEM analyses also revealed the presence of typical phagophore structures reminding those previously depicted in [43] (see blue arrowhead in Fig. 4a panel viii) and numerous intracellular compartments, containing engulfed parts of the endoplasmic reticulum, filamentous and amorphous dense structures within their lumen (see yellow arrowheads in Fig. 4a panels v, vi, x and xi). Mitochondria were also observed in close contact with the accumulated ER (see red arrows in Fig. 4a panels v to ix).
To better characterize these compartments, we initiated confocal immunofluorescence microscopy experiments on HEK293T cells co-expressing Flag-WT-USP19 and HA-TDP-43-K263E using anti-Flag (to detect USP19), anti-HA (to detect the mutant TDP-43) and anti-LC3 (to visualize endogenous LC3 positive compartments such as phagophores, autophagosomes or amphisomes). Three kinds of structures were observed: the first one corresponds to a compact structure where small inclusions/punctuated fine scattered patterns of TDP-43 are embedded and colocalized with USP19 and LC3 (see arrow and magnification in Fig. S7a,b). The second structure corresponds to TDP-43 punctuated fine scattered patterns surrounded by LC3, suggesting that TDP-43 is engulfed in an LC3-positive compartment (see arrow and magnification in Fig. S7c). The last one is an hybrid structure, where TDP-43 aggregates were embedded with LC3 and USP19 and surrounded by LC3 (see arrow and magnification in Fig. S7d). In conclusion, this set of data, correlated with the EM investigations, suggests that TDP-43, is detected with LC3 and/or USP19 positive intracellular structures reminding autophagic compartments.
Interestingly, compact electron dense structures (CEDS) were visible in TDP-43-K263E expressing cells but not in non-transfected cells (white asterisks in Fig. 4a panel ii), probably corresponding to TDP-43 aggregates. Bigger CEDS were also observed when TDP-43-K263E was co-expressed with USP19-ΔTM (white arrows in panel iii of Fig. 4a). These CEDS do probably not correspond to ER structures or USP19-ΔTM aggregates, since KDEL or Flag signal accumulations were not detected in cells co-expressing TDP-43-K263E and USP19-ΔTM (Fig. S1a, b).
We further investigated the distribution of TDP-43 in USP19-WT co-expressing cells using IEM. The data presented in Fig. 5a showed TDP-43 labeling near the accumulated ER (red arrows in panel i), within the lumen of intracellular compartments most likely corresponding to the aggregates observed in our fractionation gradients (red arrowheads in panels ii and iii), and in close contact with engulfed ER structures (red arrows in panels iii, iv) or trapped into electron dense amorphous structures (white arrowheads in panel iv).
Fig. 5
TDP-43-K263E colocalizes with USP19-WT and the KDEL-ER marker and coimmunoprecipitates with USP19-WT. a TDP-43 IEM of HEK293T TDP-43-K263E + Flag-WT-USP19 co-expressing cells. Red arrows indicate the TDP-43 gold particles in close contacts with dilated ER structures in the cytoplasm (panel i) or inside autophagic/endosomal compartments (panels iii and iv). Red arrowheads correspond to free TDP-43 aggregates in intracellular compartments (panel ii). White arrowheads correspond to TDP-43 labelling embedded in amorphous structures present in intracellular autophagic/endosomal compartments (panel iv). b Confocal immunofluorescence imaging of TDP-43-K263E + Flag-USP19-WT co-expressing cells. Co-expressing cells were labelled with antibodies directed against the ER KDEL marker (red), the Flag-USP19-WT (magenta), the HA for TDP-43-K263E (green) and the DAPI (blue) for nuclear staining. Scale bar is 10 μm. c ROI 1–2 are depicted in the merge panel. The plot profiles of the ER-KDEL marker (red) with the Flag-USP19-WT (magenta) and the HA-TDP-43-K263E (green) colocalizations (ROI1 and 2) along the ROI lines were constructed and analyzed using Image J software. White arrowheads show other colocalized signals d TDP-43 coimmunoprecipitates with Flag-USP19-WT. Left panel: co-immunoprecipitation experiment was realized on cell lysates from TDP-43-K263E and Flag-USP19-WT co-expressing cells using mouse antibodies directed against the Flag epitope or an irrelevant SARS-CoV2 IgG antibody as negative control. Right panel: Western blotting of HA-TDP-43-K263E and Flag-USP19-WT co-expressing cell lysates (input) using antibodies directed against Flag (for USP19), HA (for TDP-43) and GAPDH
IEM experiments on TDP-43-K263E and USP19-ΔTM co-expressing cells revealed TDP-43 labeling primarily associated with the CEDS and less outside these structures (red arrows in Fig. S8), thus suggesting that part of TDP-43 could be trapped within big CEDS.
Confocal immunofluorescence imaging of cells co-expressing HA-TDP-43-K263E and Flag-USP19-WT confirmed the colocalization of Flag-USP19-WT with both the ER-KDEL marker and HA-TDP-43-K263E (Fig. 5b, c).
To investigate potential physical interactions between Flag-USP19-WT and HA-TDP-43-K263E, anti-Flag immunoprecipitations experiments were conducted on lysates from cells co-expressing Flag-USP19-WT and HA-TDP-43-K263E. The data presented in Fig. 5d show that Flag-USP19-WT coimmunoprecipitates with HA-TDP-43-K263E, while no interaction was observed with a control antibody.
Overall, the physical interaction between USP19 and mutant TDP-43, along with their localization in various intracellular compartments including LC3 positive compartments, further supports the direct involvement of USP19 in the cellular handling and potential secretion of misfolded-TDP-43 species.
Role of Early autophagic and late endosomal compartments in the TDP-43-K263E secretion mediated by USP19To better characterize the cellular mechanisms by which TDP-43-K263E is secreted upon USP19-WT expression, the release of TDP-43 was explored through kinetic studies to pinpoint the onset of misfolded TDP-43 secretion. For this purpose, HEK293T cells were co-transfected with constructs as described above, and the secretion of misfolded TDP-43 was monitored by FTA ranging from 24 to 40 h post-transfection. This approach allowed us to identify the initial time point at which TDP-43 secretion becomes detectable, providing valuable insights into the temporal dynamics of this process. The data presented in Fig. S9 revealed that misfolded TDP-43 secretion becomes significant 27 h after transfection and gradually increases, reaching its peak of release at 40 h. Western blotting analyses of corresponding cell lysates revealed an increase of LC3-II protein levels compared to control (co-expression of TDP-43-K263E with USP19–ΔTM), correlating with our EM and confocal immunofluorescence data as well as with previously findings indicating that USP19 is a positive regulator of autophagy [44].
To explore the implication of the autophagic pathway in USP19-mediated secretion of misfolded TDP-43, we examined the effects of pharmacological inhibitors and small interfering RNAs (siRNAs) targeting host factors involved in intracellular trafficking and/or the biogenesis and fate of autophagic compartments (Fig. 6a).
Fig. 6
Early autophagic and endosomal compartments are involved in the TDP-43-K263E secretion mediated by USP19-WT. a Schematic representation of cellular trafficking and inhibition strategies (pharmacological in blue and siRNAs in red) used in this study to identify potential pathways involved in the TDP-43-K263E secretion mediated by USP19. b-f Evaluation of the TDP-43-K263E secretion mediated by USP19 by FTA in presence of siRNAs control (CT) or siRNAs directed against ATG7, RAB11A, HRS/HGS, RAB8A or RAB27A. Upper panel (secretion/conditioned media): nitrocellulose membranes were probed with an antibody directed against TDP-43. Lower panel (cell expression): Western blotting of cell lysates from siRNAs CT/targets-treated co-expressing cells using antibodies directed against ATG7, RAB11A, HRS/HGS, RAB8, RAB27A or Flag (for USP19), TDP-43, and GAPDH for loading control. Quantification of secreted TDP-43-K263E mediated by USP19-WT in presence of siRNAs CT and targets ATG7, RAB11A, HRS/HGS, RAB8A and RAB27A are depicted in the right panels of each condition. Data represent mean ± SEM, n = 4 to 6 experiments. Significance was assessed by a Mann–Whitney U test (*p < 0.05)
The VPS34 protein (class III phosphatidylinositol 3 kinase PI3K) plays a crucial role in the early steps of autophagosome biogenesis, and its inactivation impairs autophagosome formation[45, 46]. Pharmacological inhibitor of VPS34, Spautin-1 and LY294002, significantly reduced the secretion of misfolded TDP-43 compared to the DMSO control (Fig. S10a, b). Inhibition of VPS34 was confirmed by Western blotting showing the inhibition of S6 phosphorylation, a downstream effector of VPS34.
To confirm the implication of autophagosomes, ATG7, a key factor involved in early autophagosome biogenesis, was silenced using ATG7 siRNAs. The data presented in Fig. 6b revealed that ATG7 siRNA reduced the secretion of aggregated TDP-43. Similarly, RAB11A has also been found to be required for the recruitment and assembly of the autophagic machinery [47]. Consistently, when we conducted a RAB11A siRNA, we observed a significant reduction of USP19-mediated TDP-43-K263E secretion (Fig. 6c).
Altogether, these findings indicate that autophagosomes are involved in USP19-mediated secretion of misfolded TDP-43.
Autophagosomes can fuse with late endosomal compartments to form hybrid amphisomal compartments, and the endosomal sorting complex required for transport ESCRT-0 HRS/HGS component plays a key role in late endosomes and amphisomes biogenesis [48, 49]. We found that silencing HRS/HGS expression efficiently decreased USP19-mediated TDP-43-K263E secretion indicating the involvement of the late endosomal/amphisome compartments (Fig. 6d).
Autophagosomes or amphisomes can fuse with lysosomes to create hybrid structures called autophagolysosomes, where protein enzymatic degradation occurs. To determine if lysos
Comments (0)