
Remember me

A list of all of the bacterial strains, plasmids and oligonucleotides used in this study is provided in Supplementary Tables 1–4. LB and LB agar were used for cloning and maintenance of strains. For selection in B. subtilis, media were supplemented with the following antibiotics when required: spectinomycin (80 µg ml−1), chloramphenicol (5 µg ml−1), kanamycin (10 µg ml−1) and tetracycline (10 µg ml−1). The combination of lincomycin (12.5 µg ml−1) and erythromycin (0.5 µg ml−1) was used to select for macrolide–lincosamide–streptogramin (MLS) resistance. Carbenicillin (100 µg ml−1) was used for selection in E. coli.
B. subtilis (p)ppGpp biosynthesis mutants were constructed by transformations of integration plasmids containing an I-sceI endonuclease cut site and regions of homology upstream and downstream of specific synthetase genes (pJW300 for ∆sasB; pJW370 for sasBF42A; pJW306 for ∆sasA; and pJW371 for relD264G) followed by transformation of pSS4332 for marker-less recombination50. Successful recombination was verified by PCR. For the construction of the (p)ppGpp0 mutant, ∆sasA ∆sasB cells were transformed with ∆rel::mls PCR product from genomic DNA using the oligos oJW902 and oJW903 followed by MLS resistance selection26. Construction of integration plasmid for sasBF42A was performed using site-directed mutagenesis of pJW370 by PCR using the oligos oJW2309 and oJW2310.
The (p)ppGpp0gmkQ110R mutant was obtained from isolating suppressor mutants from (p)ppGpp0 cells by plating on S7 minimal medium plates containing 1% glucose. The surviving colonies were plated on S7 minimal medium plates containing 0.5% casamino acids and 0.5 mM 8-azaguanine, or S7 minimal medium plates containing 0.5% casamino acids and 0.1 mM guanosine26 to differentiate between mutants with mutations in hprT or gmk. Colonies that can grow on guanosine but not 8-azaguanine were sequenced to identify the mutant gmk allele. Whole-genome sequencing was performed to confirm that gmkQ110R is the only mutation in the strain.
The guaBdown mutant in the (p)ppGpp+ background was constructed by transformation of pJW30526, which replaces the chromosomal copy of guaB with an isopropyl-β-d-1-thiogalactopyranoside (IPTG)-inducible copy of guaB (Pspac-guaB). This enables controllable guaB expression using IPTG during strain construction and growth to avoid generation of suppressors.
B. subtilis deletion mutants were constructed by serial transformation of PCR products from the B. subtilis knockout collection (BGSC, Gross laboratory)51. Where required, the lox-site-flanked ermR or kanR cassette was removed using pDR244-cre followed by selection for the loss of MLS or Kan resistance.
Construction of PlowGTP fluorescence reporters was done by fusion of PCR products containing the PlowGTP promoter (primers oJW1935 and oJW1936) with coding regions of fluorescence proteins (primers oJW1995 and oJW1996 (for GFP) or oJW2805 and oJW2806 (for mCherry)) using ligase cycling reaction (LCR)52. The promoter–fluorescence protein gene fusions were cloned into the pDR110 backbone flanked by amyE without the Pspank promoter for subsequent transformation.
For the construction of the PrrnBP1-GFPns (unstable GFP sequence described previously53) reporter, DNA fragments of PrrnBP1 (primers oJW2083 and oJW2084), GFPns (primers oJW1995 and oJW2020), the flanking regions of lacA (primers oJW1990 and oJW2414, and oJW2413 and oJW2082) and lox-site-flanked ermR cassette (primers oJW2133 and oJW2134) were amplified by PCR using synthetic oligonucleotides or genome DNA. The resulting PCR products were fused by LCR followed by amplification using PCR to generate the linear recombination fragment of lacA::PrrnBP1-GFPns-lox-ermR-lox for transformation.
For the construction of PsasA-mCh reporter, DNA fragments of PsasA (primers oJW3099 and oJW3079), mCh (primers oJW2805 and oJW2806), the flanking regions of lacA (primers oJW1990 and oJW2414, and oJW2413 and oJW2082) and lox-site-flanked ermR cassette (primers oJW2133/oJW2134) were amplified by PCR using synthetic oligonucleotides or genome DNA. The resulting PCR products were fused by LCR followed by amplification using PCR to generate the linear recombination fragment of lacA::PsasA-mCh-lox-ermR-lox for transformation.
For the construction of Pveg-GFP reporter, DNA fragments of Pveg (primers oJW3928 and oJW2806), GFP (primers oJW1995 and oJW1996), the flanking regions of lacA (primers oJW1990 and oJW2414, and oJW2413 and oJW2082) and lox-site-flanked ermR cassette (primers oJW2133 and oJW2134) were amplified by PCR using synthetic oligonucleotides or genome DNA. The resulting PCR products were fused by LCR followed by amplification using PCR to generate the linear recombination fragment of lacA:: Pveg-GFP-lox-ermR-lox for transformation.
Removal of the lox-site flanked ermR cassette was done by transformation with pDR244-cre and selecting for the loss of MLS resistance. All of the mutants and constructs were verified by DNA sequencing.
Growth conditionsB. subtilis strains were grown in S7 defined medium54; MOPS was used at 50 mM rather than 100 mM, supplemented with 0.1% glutamate, 1% glucose and 0.5% casamino acids. Growth of the YB886 strain background was supplemented with 20 µg ml−1 tryptophan and 50 µg ml−1 methionine. For growth in minimal medium, both glutamate and casamino acids were replaced with 200 µg ml−1l-isoleucine, 200 µg ml−1l-leucine and 200 µg ml−1l-valine, and 1% carbon sources were used as indicated.
Routinely, cells from young colonies on overnight LB-agar plates at 37 °C (<12 h) were inoculated into growth medium and then grown at 37 °C with 250 rpm shaking. Cultures in logarithmic phase (optical density at 600 nm (OD600) of around 0.1–0.3) were treated with antibiotics or inducers, including arginine hydroxamate (RHX, 0.5 mg ml−1), carbonyl cyanide m-chlorophenyl hydrazone (CCCP, 5 µM), sodium azide (NaN3, 4 mM) or arsenate (2.5 mM). IPTG was added to a final concentration of 0.5 mM to induce guaB expression from an IPTG-inducible promoter (Pspac), while depletion of guaB expression was done by omitting IPTG in the growth medium.
The following inducers and concentrations were used unless otherwise specified: RHX, 0.5 mg ml−1; CCCP, 5 μM; NaN3, 4 mM; arsenate, 2.5 mM; carbenicillin, 0.5 μg ml−1 (0.5× MIC) or 100 μg ml−1 (200× MIC); bacitracin, 64 μg ml−1 (0.5× MIC) or 384 μg ml−1 (3× MIC); ciprofloxacin, 0.1 μg ml−1 (0.5× MIC) or 4 μg ml−1 (20× MIC); kanamycin, 0.625 μg ml−1 (0.3× MIC) or 8 μg ml−1 (8× MIC); vancomycin, 0.1 μg ml−1 (0.5× MIC) or 4 μg ml−1 (20× MIC) along with the non-induction controls.
MIC determinationMICs for chloramphenicol, tetracycline, kanamycin, ciprofloxacin, norfloxacin, rifampicin, bacitracin and vancomycin were determined using the microdilution method55. Logarithmic-phase cells were back-diluted to a final titre of around 5 × 105 colony-forming units (CFU) per ml into 96-well plates containing twofold serial dilutions of respective antibiotics in S750 medium with 0.1% glutamate, 1% glucose and 0.5% casamino acids. After incubation for 16–20 h at 37 °C with 250 rpm shaking, the MIC was determined as the lowest drug concentration that prevented visible growth.
Bacterial growth measurementFor growth measurement, fresh colonies of B. subtilis strains on LB agar were resuspended into different growth media as specified and diluted to OD600 ≈ 0.005 in 96-well plates. Growth was monitored by OD600 at 37 °C under shaking in the Synergy2 microplate reader (BioTek). Doubling times were estimated by fitting the growth data to the exponential growth curve using a custom Python script.
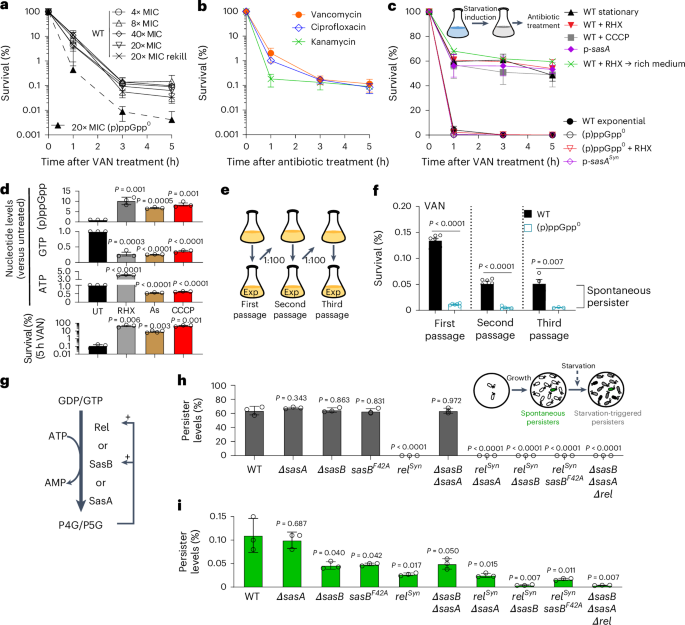
Persister assayTo prepare exponentially growing B. subtilis populations, cells from young colonies on overnight LB-agar plates at 37 °C (<12 h) were inoculated into S750 medium with 0.1% glutamate, 1% glucose and 0.5% casamino acids, and grown to OD600 ≈ 0.1–0.3 at 37 °C, 250 rpm. For growth in minimal medium, both glutamate and casamino acids were replaced with 200 µg ml−1l-isoleucine, 200 µg ml−1l-leucine and 200 µg ml−1l-valine, and 1% carbon sources were used as indicated. Treatments with bactericidal antibiotics were performed at the following concentrations: ciprofloxacin, 4 μg ml−1 (20× MIC); vancomycin, 4 μg ml−1 (20× MIC); kanamycin, 8 μg ml−1 (8× MIC); and bacitracin, 384 μg ml−1 (3× MIC). To determine cell viability, culture aliquots were taken at T = 0 and at designated times after treatment, serially diluted and plated onto LB agar. Plates were incubated at 37 °C overnight. The viability at different timepoints, determined as the CFU per ml and relative survival (versus T0), was calculated.
For experiments involving pre-induction of cells with (p)ppGpp-inducing agents, cells were grown to OD600 ≈ 0.1 and divided into two cultures: one containing the inducing agent (RHX, 0.5 mg ml−1; bacitracin, 64 μg ml−1; CCCP, 5 μM; NaN3, 4 mM; arsenate, 12.5 mM) and other as a non-induction control. The cultures were grown for an additional 30 min under the same conditions (T = 0.5 h) and subjected to the persister assay, as described above.
In the case of measuring spontaneous persistence, we defined spontaneous persisters as those that are generated during growth in a non-stressed condition1,3. To achieve this, early-exponentially growing cultures were 1:100 back-diluted into fresh medium and regrown to exponential phase (OD600 ≈ 0.1–0.3) for one or two rounds followed by antibiotic treatment1.
Estimation of antibiotic toleranceFor the estimation of population tolerance, we used the MDK99 (ref. 1). To exclude the contribution of persisters to population tolerance, we determined the level of persisters in the biphasic killing curve and subtracted this subpopulation from the bulk population. The MDK99 of the population was estimated from the logarithmic killing phase in the killing curve.
Measurement of intracellular nucleotides using TLCTo measure intracellular nucleotides, cells were first collected from overnight plates, back-diluted to OD600 = 0.005 and grown in low-phosphate (0.1× phosphate, 0.5 mM) S750 medium with 0.1% glutamate, 1% glucose and 0.5% casamino acids. Once cultures reached OD600 ≈ 0.05, 1 ml cells was labelled with 50 µCi of 32P orthophosphate (900 mCi mmol−1; Perkin Elmer) for 2–3 generations before treatment or sampling. At OD600 ≈ 0.15, RHX, CCCP or arsenate were added to the cultures and the samples were collected at regular timepoints for nucleotide extraction. Nucleotides were extracted by incubating 100 µl cells with 20 µl of 2 M formic acid on ice for at least 20 min. The samples were spotted onto PEI cellulose TLC plates (Millipore, 1055790001) and resolved in 1.5 M or 0.85 M potassium phosphate monobasic (KH2PO4, pH 3.4) buffer to separate (p)ppGpp or GTP, respectively. TLC plates were exposed on storage phosphor screens (GE Healthcare) and scanned on the Typhoon imager (GE Healthcare).
Measurement of intracellular nucleotides by LC–MSLC–MS quantification of nucleotides was performed as described previously21. Cells were grown in S750 medium supplemented with 20 amino acids21 to OD600 ≈ 0.3 at 37 °C, 250 rpm before collection. Then, 25 ml of cultures was sampled and filtered through PTFE membrane (Sartorius, 14555419). For experiments involving bacitracin induction, cells were collected before and after 0.25× MIC (sublethal) or 1.25× MIC (lethal) bacitracin treatment for 30 min. Membranes with cell pellets were submerged in 3 ml extraction solvent mix (on ice 50:50 (v/v), chloroform:water) to quench metabolism, lyse the cells and extract metabolites. Mixtures of cell extracts were centrifuged at 5,000g for 10 min to remove organic phase and then centrifuged at 20,000g for 10 min to remove cell debris. The samples were analysed using an HPLC–MS system, consisting of a Vanquish UHPLC system linked through electrospray ionization (ESI, negative mode) to the Q Exactive Orbitrap mass spectrometer (Thermo Fisher Scientific) operated in full-scan mode to detect targeted metabolites based on their accurate masses. LC was performed on the Acquity UPLC BEH C18 column (1.7 μm, 2.1 × 100 mm; Waters). The total run time was 30 min with a flow rate of 0.2 ml min−1, using solvent A as denoted above and acetonitrile as solvent B. The gradient was as follows: 0 min, 5% solvent B; 2.5 min, 5% solvent B; 19 min, 100% solvent B; 23.5 min, 100% solvent B; 24 min, 5% solvent B; 30 min, 5% solvent B. Quantification of metabolites was performed using MAVEN software56 and normalized to the OD600 at the time of cell collection.
For LC–MS analysis of FACS-sorted cells, membranes with filtered cells (about 8 × 106 cells) were submerged in 1.5 ml extraction solvent mix (methanol:acetonitrile:H2O, 40:40:20) to quench metabolism, lyse the cells and extract metabolites. The cell extract was centrifuged at 21,000g for 10 min at 4 °C, and 1 ml of supernatant was then transferred to a new microcentrifuge tube and dried completely with the SpeedVac. Dried metabolites were then resuspended in 100 μl solvent A (97:3 (v/v) water:methanol, 10 mM tributylamine ~pH 8.2–8.5 adjusted with ~9 mM acetic acid). The samples were analysed using an HPLC-MS system consisting of a Vanquish UHPLC system linked through heated electrospray ionization (ESI, negative mode) to a hybrid quadrupole high-resolution mass spectrometer (Q-Exactive Orbitrap, Thermo Fisher Scientific) operated in full-scan selected ion monitoring (MS-SIM) mode to detect targeted metabolites on the basis of their accurate masses. MS parameters were set to a resolution of 140,000, an automatic gain control (AGC) of 3 × 106, a maximum injection time of 100 ms and a scan range of 400–1,000 m/z. Only ions with a retention time of 10–15 min were scanned by MS. LC was performed on the Acquity UPLC BEH C18 column (1.7 μm, 2.1 × 100 mm; Waters). The total run time was 30 min with a flow rate of 0.2 ml min−1, using solvent A and 100% acetonitrile as solvent B. The gradient was as follows: 0 min, 5% B; 2.5 min, 5% B; 19 min, 100% B; 23.5 min, 100% B; 24 min, 5% B; and 30 min, 5% B. Raw output data from the MS was converted to mzXML format using custom software. Quantification of metabolites was performed using MAVEN software56 and normalized to an internal standard of six most represented nucleotides detected in the sample.
Fluorescence microscopyTo monitor (p)ppGpp induction using fluorescence reporters, cells were grown to OD600 ≈ 0.1–0.3 followed by 30 min induction with the following inducers at concentrations listed below unless otherwise specified: RHX, 0.5 mg ml−1; CCCP, 5 μM; NaN3, 4 mM; arsenate, 2.5 mM; carbenicillin, 0.5 μg ml−1 (0.5× MIC) or 100 μg ml−1 (200× MIC); bacitracin, 64 μg ml−1 (0.5× MIC) or 384 μg ml−1 (3× MIC); ciprofloxacin, 0.1 μg ml−1 (0.5× MIC) or 4 μg ml−1 (20× MIC); kanamycin, 0.625 μg ml−1 (0.3× MIC) or 8 μg ml−1 (4× MIC); vancomycin, 0.1 μg ml−1 (0.5× MIC) or 4 μg ml−1 (20× MIC), along with the non-induction controls.
All of the imaging samples were spotted onto 1.5% agarose pads made with the same growth medium, and immediately imaged on the Olympus IX-83 inverted microscope (Olympus) using a ×60 phase-contrast objective with fluorescence filters (excitation: 470/20 nm, dichroic mirror: 485 nm, emission: 515/50 nm for GFP; excitation: 575/20 nm, dichroic mirror: 595 nm, emission: 645/90 nm for mCherry or propidium iodide; and excitation: 427/10 nm, dichroic mirror: 595 nm, emission: 472/30 nm for Sytox Blue). Metamorph Advanced (v.7.8.3.0) (Molecular Devices) was used for microscopy data collection. Single-cell time-lapse imaging was performed at 15 min intervals for each field at 37 °C using a temperature-controlled imaging chamber (Tokai Hit) coupled to an automatic stage and the microscope control as described previously57. The measurement was generally over the course from the birth of the cell until the time lapse stopped owing to crowding of the microcolony or, in rare cases, severe drifting of focus. When comparing phenotypes between strains, both strains were imaged in parallel on the same imaging dish using the same microscope with same settings. For imaging persister survival in time-lapse experiments, final concentrations of 5 μg ml−1 carbenicillin (10× MIC) and 200 nM Sytox Blue or propidium iodide (Molecular Probes) were applied to the agarose pads at designated times. To remove carbenicillin after treatment, 5 U ml−1 final concentration of penicillinase (Sigma-Aldrich, P0389) was applied. Strains without the fluorescence reporters were used for autofluorescence measurement.
Biofilm growth and imagingB. subtilis biofilms were grown on a custom microfluidic device fabricated with polydimethylsiloxane. The device contains a central chamber connected to inlet and outlet media channels, allowing for constant medium flow through the central chamber. A semipermeable dialysis membrane was fixed on top of the central chamber to provide a platform for biofilm growth. This setup allows diffusion of nutrients or small molecules from the medium flow underneath to support biofilm growth on the membrane and allows subsequent treatment with antibiotics. To grow B. subtilis biofilms, 1 µl of early exponential phase culture (OD600 ≈ 0.05) was applied onto the membrane and grown at room temperature (25 °C) for 24 h under a constant flow of S750 medium supplemented with 0.5% glutamate and 0.5% glycerol.
Imaging of biofilms was performed with the IXplore SpinSR confocal imaging microscope (Olympus) using a ×20 phase-contrast objective with fluorescence filters (488 nm laser with 510–550 nm emission filter for GFP; 561 nm laser with 575–625 nm emission filter for mCherry). Biofilms at 24 h after inoculation were imaged before and after switching to the same growth medium containing 4 µg ml−1 vancomycin. In total, 51 stacks at 1 µm intervals were taken for each timepoint. Images were projected along the z axis from the top of the biofilm using maximum-intensity projection using cellSense Dimension v.2.2 (Olympus). For quantitation of reporter signals within the biofilm, we sampled around four random regions within the centre of the biofilm and measured their GFP and mCherry intensities. The background fluorescence was measured from regions without biofilm and used for background subtraction.
Flow cytometry and cell sortingFlow cytometry was performed at the UWCCC flow cytometry core. To prepare samples for flow cytometry, cells from young colonies on overnight LB-agar plates at 37 °C (<12 h) were inoculated into and grown in S750 medium with 0.1% glutamate, 1% glucose and 0.5% casamino acids to OD600 ≈ 0.1–0.3 at 37 °C, 250 rpm. For growth in minimal medium, both glutamate and casamino acids were omitted and 1% carbon sources were used as indicated. Cells were immediately fixed with 0.4% paraformaldehyde for 15 min at room temperature, washed three times with 1× PBS and kept at 4 °C until analysis. Fixation was verified by viability plating and microscopy. Flow cytometry analysis was performed using the BD LSRFortessa flow cytometer (BD Biosciences) with a 70 µm nozzle. BD FACSDiva v.8.0.2 was used for data collection. Cell populations were detected using both forward and side scatter (FSC and SSC). Single-cell fluorescence was measured using the 488 nm laser and detection filters for GFP (530/30 nm, 505LP dichroic filter). Autofluorescence was measured by analysing parental strains without the fluorescence reporter and subtracted from the raw reporter fluorescence. Approximately 1.5 million events were measured for each sample. For the determination of antibiotic-induced persistence, the frequency of low-GTP cells after antibiotic induction was subtracted from the frequencies before induction.
FACS was performed at the UWCCC flow cytometry core. To prepare samples for cell sorting, cells were collected from young colonies on overnight LB-agar plates at 37 °C (<12 h) and grown in S750 medium with 0.1% glutamate, 1% glucose and 0.5% casamino acids to OD600 ≈ 0.3 at 37 °C, 250 rpm. FACS analysis was performed using the BD FACSAria cell sorter (BD Biosciences) with a 70 µm nozzle at room temperature using the 488 nm laser and 530/30 nm detection filters for GFP, and the 561 nm laser and 610/20 nm detection filters for mCherry. BD FACSDiva v.8.0.2 was used for data collection. Autofluorescent cells were eliminated by gating using an isogenic strain without the fluorescent reporters. At least 1,000 cells were obtained from the rarest gate for each sample. Cell recovery rate was estimated to be >90% based on viability counting on LB plates. For antibiotic treatment, cells were directly sorted into tubes containing 4× MIC of vancomycin followed by shaking at 37 °C. Aliquots were taken at different times for serial dilution and plating to measure survival by colony counting. The number of cells before treatment (T0) was measured using the cell sorter.
For FACS-sorting of cells for LC–MS analysis, cells containing the PlowGTP reporter were grown in S750 medium with 0.1% glutamate, 1% glucose and 0.5% casamino acids to either exponential (OD600 ≈ 0.2) or stationary (OD600 ≈ 4.0) phase. Both populations were mixed at a 10:1 ratio, and immediately FACS-sorted into low-fluorescence or high-fluorescence fractions. Autofluorescent cells were eliminated by gating using an isogenic strain without the fluorescent reporters. Approximately 8 × 106 cells were sorted and filtered on the PTFE membrane (Sartorius, 14555419) to remove the sheath fluid. Membranes with filtered cells were then subjected to metabolite extraction and LC–MS analysis. CFU count analysis was performed from a small aliquot of the sorted fractions to account for potential variations in the number of cells sorted between fractions.
Tn-seqThe B. subtilis 168 transposon mutant library was provided by the Grossman laboratory58. Construction of the library was performed as follows. In brief, in vitro transposition of B. subtilis 168 genomic DNA (gDNA) with magellen6x transposon was performed by mixing 1.3 µg pCJ41 (containing magellen6x transposon), 34 ng purified MarC9 transposase, 5 µg B. subtilis gDNA, 10 µl 2× buffer A (41 mM HEPES pH 7.9, 19% glycerol, 187 mM NaCl, 19 mM MgCl2, 476 µg ml−1 BSA and 3.8 mM DTT) into a 20 µl reaction in vitro and incubated overnight at 30 °C. The transposed DNA was precipitated and resuspended in 2 µl 10× buffer B (500 mM Tris-Cl pH 7.8, 100 mM MgCl2, 10 mM DTT), 2 µl 1 mg ml−1 BSA and 11 µl H2O followed by 4 h incubation at 37 °C. After incubation, 4 µl of 2.5 mM dNTPs and 1 µl of 3U µl−1 T4 DNA polymerase were added to the DNA and further incubated for 20 min at 12 °C, followed by heat-inactivation at 75 °C for 15 min. Next, 0.2 µl 2.6 mM NAD and 1 µl of 10 U µl−1E. coli DNA ligase were added and the reaction was incubated overnight at 16 °C. The resulting in vitro transposed and repaired gDNA was transformed into B. subtilis 168 and plated onto LB agar containing spectinomycin and incubated overnight. Colonies containing the transposon were washed off and pooled into a single library. The library was estimated to contain around 50,000 unique transposon inserts across the genome.
For the selection experiment with the transposon library, an aliquot of the library was inoculated and grown in S750 medium with 0.1% glutamate, 1% glucose and 0.5% casamino acids supplemented with tryptophan (20 µg ml−1) at 37 °C, 250 rpm. At OD600 ≈ 0.1–0.3, the cultures were treated with 20× MIC vancomycin or ciprofloxacin. Cultures before and after antibiotic treatment were plated onto LB plates and recovered after incubation for around 14 h at 37 °C. Around 650,000 colonies from each sample were pooled and snap-frozen for gDNA extraction and sequencing library preparation.
Preparation of the sequencing library was performed as previously described59. Frozen cell pellets were resuspended in 500 µl lysis buffer with lysozyme and RNase A (20 mM Tris-HCl pH 7.5, 50 mM EDTA, 100 mM NaCl, 2 mg ml−1 lysozyme, 120 µg ml−1 RNase A) and incubated at 37 °C for 20–30 min. Next, the incubated cell lysate was mixed with 60 µl 10% N-lauroylsarkosine and further incubated at 37 °C for 15 min. gDNA was purified using 600 μl phenol, then 600 µl phenol:chloroform:isoamyl alcohol (25:24:1) and finally 600 µl pure chloroform. DNA in the aqueous phase was precipitated using 1/10 volumes of 3 M NaOAc and 2 volumes of 100% ethanol. The DNA pellet was washed with 70% ethanol, air-dried on the bench and resuspended in 10 mM Tris-HCl, pH 8.5 and stored at 4 °C. For each sample, 6 µg of DNA was used for MmeI digestion in 200 µl (6 µg gDNA, 6 µl MmeI (2,000 U ml−1, NEB), 0.5 µl 32 mM S-adenosylmethionine, 20 µl NEB CutSmart Buffer and double-distilled H2O up to 200 µl). DNA was digested for 2.5 h at 37 °C, after which 2 µl calf intestinal phosphatase (10,000 U ml−1, NEB) was added to the digest and the sample was incubated for 1 h at 37 °C. Digested gDNA was extracted with 200 µl phenol:chloroform:isoamyl alcohol (25:24:1) followed by 200 µl of pure chloroform. DNA in the aqueous phase was first mixed with 1/10 volume of 3 M NaOAc and 67 ng ml−1 glycogen, and then 2.5 volumes of 100% ethanol. The tubes were then placed at −80 °C for 20 min and then centrifuged at maximum speed for 15 min at 4 °C. Precipitated DNA was washed with 150 µl 70% ethanol twice at room temperature, air-dried and resuspended in 15 µl of double-distilled H2O.
For annealing of the DNA adaptor, 20 µl of 100 µM synthesized oligos (IDT) were mixed with 1 µl of 41 mM Tris-HCl pH 8.0 (final concentration of adaptor, 50 µM in 1 mM Tris-HCl pH 8.0). Oligos were annealed by heat denaturation (95 °C for 5 min) and stepwise cool-down (94 °C for 45 s then repeat with −0.3 °C per cycle for 250 cycles, then hold at 15 °C) using a PCR machine. Annealed adaptors were diluted to 3.3 µM in double-distilled H2O and stored at −20 °C.
For adaptor ligation, 5 µl of digested DNA was mixed with 1 µl of 3.3 µM DNA adaptor, 1 µl of 10× T4 DNA ligase buffer (NEB), 1 µl of T4 DNA ligase (400,000 U ml−1, NEB) and 2 µl double-distilled H2O. The ligation mix was incubated overnight at 16 °C in a PCR machine.
Amplification of the adaptor-ligated DNA library was performed using barcoded primers and Phusion high-fidelity DNA polymerase (NEB) for 18 cycles according to the provided instructions. The PCR products were mixed in equal amounts, purified by size exclusion and submitted for sequencing using the Illumina sequencing primer (5′-ACACTCTTTCCCTACACGACGCTCTTCCGATCT-3′). Deep sequencing was performed on the Illumina HiSeq 2500 (Illumina) system by the University of Michigan DNA Sequencing Core. Analysis of sequencing data was performed using a custom Python script and mapped to the B. subtilis 168 reference genome (NCBI: NC_000964.3). Visual inspection of transposon insertion profiles was done using GenomeBrowse (Golden Helix).
Expression and purification of B. subtilis RelThe plasmid for Rel purification was constructed as follows. The B. subtilis rel coding sequence was PCR amplified from NCIB3610 genomic DNA using the primers oJW3196 and oJW3197. The pE-SUMO expression vector was amplified using the primers oJW3194 and oJW3195. The PCR products were assembled to generate pJW753 by Golden Gate assembly (New England BioLabs). Plasmids were verified by DNA sequencing.
To express His6–SUMO–Rel, fresh transformants of E. coli BL21 carrying pE-SUMO-rel were grown in LB at 37 °C to OD600 ≈ 0.5, followed by 1:50 dilution into Terrific Broth and grown at 30 °C until OD600 ≈ 1.5. His6–SUMO–Rel expression was induced with 1 mM IPTG for 4 h at 30 °C. Cells were pelleted and stored at −80 °C until use. Frozen cell pellets were thawed on ice, resuspended in ice-cold lysis buffer (50 mM Tris-HCl, pH 8, 1 M NaCl, 10 mM imidazole, DNase and cOmplete protease inhibitor (Roche)), and lysed using a French press at 4 °C. The cell lysate was centrifuged at 4 °C, 16,000g for 30 min to obtain the supernatant. The filtered supernatant was injected into the ӒKTA FPLC system (GE Healthcare) and passed through the HisTrap FF column (GE Healthcare). His6–SUMO–Rel was eluted with a gradient of buffer A (50 mM Tris-HCl, pH 8, 1 M NaCl, 5% glycerol, 10 mM imidazole) and buffer C (50 mM Tris-HCl, pH 8, 1 M NaCl, 5% glycerol, 500 mM imidazole). The fractions containing the protein were pooled with 300 µl SUMO protease into Spectra/Por dialysis tubing (Spectrum), and dialysed into 50 mM Tris-HCl, 1 M NaCl, 1 mM β-mercaptoethanol and 5% glycerol overnight. Rel without the His6–SUMO tag was passed through the HisTrap FF column and then purified by size exclusion using the Superose 12 10/300 GL column (GE Healthcare) with the ӒKTA FPLC system. The fractions containing the Rel protein were pooled and measured for its concentration using the Bradford Assay (Bio-Rad). Aliquots were snap-frozen using liquid nitrogen and stored at −80 °C.
In vitro pppGpp synthesis assayIn vitro pppGpp synthesis by Rel was monitored by measuring synthesis of radiolabelled pppGpp over time. The reaction contained 236 nM B. subtilis Rel, 0.05 µM [α32P]GTP, 1 mM ATP, 50 mM NaCl and 10 mM ATP in 20 mM Tris-HCl pH 7.5, with or without 10 µM non-radioactive pppGpp. The reaction lacked manganese to avoid a potential effect of pppGpp hydrolysis. The reaction was initiated by the addition of ATP and incubated at 37 °C. At the indicated times, 10 µl of the reaction was mixed with 2 µl of 2 M formic acid and chilled on ice for 20 min to quench the reaction. Then, 1 µl samples were spotted onto PEI cellulose TLC plates (Millipore, 1055790001) and resolved in 1.5 M potassium phosphate monobasic (KH2PO4, pH 3.4) buffer to separate pppGpp. TLC plates were dried and exposed on storage phosphor screens (GE Healthcare) and scanned on the Typhoon imager (GE Healthcare).
Quantification and statistical analysisFor TLC experiments, nucleotide spots were quantified using ImageQuant v.5.0 (Molecular Dynamics). The raw intensities were corrected to the number of phosphates in the corresponding nucleotide and normalized to OD600 or ATP level before treatment (ATPT=0) for comparison between samples. For in vitro pppGpp synthesis assays, changes in pppGpp levels were normalized to T = 0.
Microscopy image analysis and cell parameters (cell area and fluorescence intensity) measurements were performed using Metamorph Advanced (v.7.8.3.0) (Molecular Devices). Background and autofluorescence were subtracted by comparing images obtained from identical strains without the fluorescence reporter. The single-cell specific growth rate (µ) at each frame was calculated using the equation \(\mu =\frac\frac\), where l is the cell length, Δl is the change in cell length and Δt is the change in time (in min). Flow-cytometry data were analysed using FlowJo X (FlowJo); an example gating strategy is shown in Supplementary Fig. 14a. Cells within a narrow range of cell sizes were gated, subgated to filter cell aggregates and then measured for their fluorescence distribution. Autofluorescence was measured using isogenic strains without the fluorescent reporters and subtracted from raw reporter signals. Gating for PlowGTP-high cells was set at fivefold or higher above mean population fluorescence. This cut-off agrees with our FACS sorting experiment in which cells with reporter fluorescence above this threshold are predominantly persisters. For the determination of antibiotic-induced persistence, the frequency of low-GTP cells after antibiotic induction was subtracted from the frequencies before induction.
Statistical information of individual experiments is included in the figure legends. n represents the number of biological replicates or number of cells for experiments involving single-cell measurements as indicated in the legends. Significance was tested using unpaired two-tailed t-test or as specified in the figure legend. Confidence intervals were calculated based on binomial distribution. Prism 7 (GraphPad) was used for statistical analysis.
Reporting summaryFurther information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Comments (0)