Liver diseaseHCC
Hepatocellular carcinoma (HCC), the predominant type of liver cancer, represents an escalating clinical challenge. Globally, HCC ranks as the fifth most prevalent cancer and the third leading cause of cancer-related mortality. According to previous gene expression profiling studies, OPN demonstrates significantly elevated expression levels in HCC, serving as a poor prognostic indicator for the disease (Wen and Jeong et al., 2016). Furthermore, studies indicate that OPN is primarily secreted by HCC cells in liver cancer, with significant expression observed in both Hep G2 cells and HCC-LM3 cells. The study discovered that HCC patients’ serum OPN levels were significantly elevated compared to those in individuals with chronic liver disease (CLD) and healthy controls. OPN levels were higher in patients with early-stage HCC than those with CLD, and OPN levels were higher in individuals with advanced HCC (Qin 2014). OPN also demonstrates high sensitivity in detecting AFP-negative hepatocellular carcinoma, suggesting its potential as a superior biomarker for early-stage HCC and as a diagnostic marker reflecting disease progression ((Shang and Plymoth et al., 2012; Wen and Jeong et al., 2016; Tan and Zhao et al., 2022). OPN levels in tissue and blood were linked to survival, stage, tumor size and liver dysfunction (Simão and Madaleno et al., 2015; Sun and Li et al., 2018). In addition, an elevation in OPN levels was also observed in primary hepatocellular carcinoma with metastasis. A study assessing the metastatic factors of HCC identified OPN as a key characteristics associated with metastasis (Sun and Dong et al., 2008). Researcher has demonstrated that vascular and bile duct invasion, the Edmonson grade system, and intrahepatic invasion are significantly associated with elevated OPN levels in tumor tissues from a substantial cohort of HCC patients (Ye and Qin et al., 2003; Qin 2014). Specifically, patients with high OPN expression demonstrate markedly poorer tumor recurrence time (TTR) compared to those with low OPN expression (Zhou and Zhou et al. 2013; Qin 2014). These researches demonstrate that OPN may outperform AFP as an HCC marker, offering valuable predictive insight even in early HCC stages (Ye and Qin et al., 2003; Zhang and Ye et al., 2006; Mack and Redman et al., 2008; Qin 2014). And higher plasma OPN levels are linked to a greater risk of tumor recurrence after liver cancer surgery.
The overexpression of OPN has been shown to enhance proliferation and metastasis both in vitro and in vivo (Wen and Jeong et al., 2016). Knockdown of SPP1 in HCC-LM3 and HepG2 cells has been shown to affect cell proliferation, migration and invasion. The biological function of OPN in promoting cancer is contingent upon CD44 and surface integrins, with a specific isoform of OPN-a demonstrating the most significant proliferative effect (Ge and Lu et al., 2013). OPN interacts with the CD44 receptor to augment hepatocyte growth factor (HGF)-induced cellular scattering and invasion, and it activates the c-Met signaling pathway to facilitate the progression of HCC (Medico and Gentile et al., 2001; Yoo and Gredler et al., 2011). Multiple signaling pathways, including MAPK, JNK, PI3K, FAK, and Src, which collectively facilitate EMT, as well as cell migration and invasion, are activated in response to the binding of secreted OPN to its receptors in HCC.
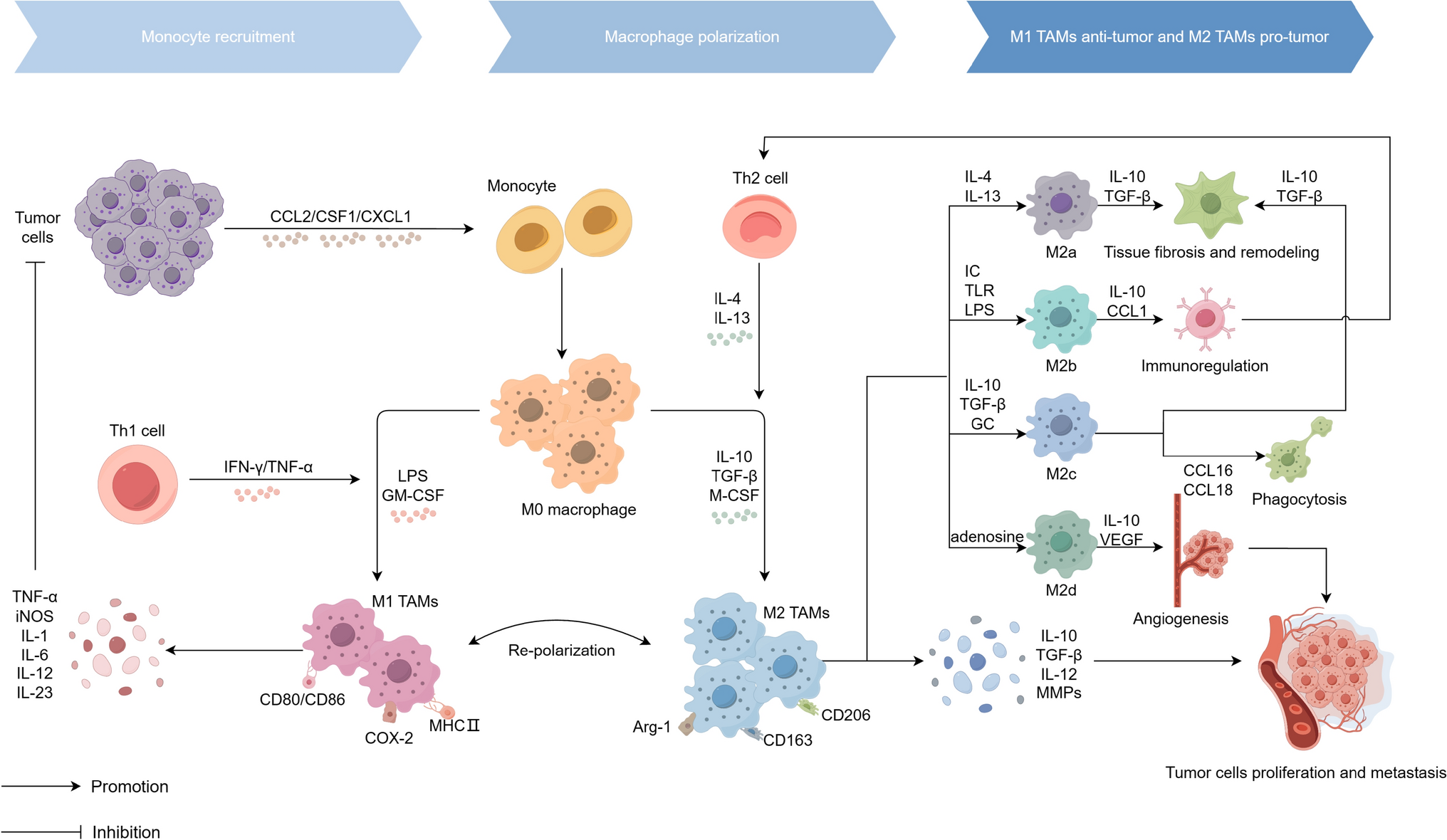
OPN play a critical role in recruiting tumor-associated macrophages tomodulate tumor progression. This process it is primarily mediated throught interactions with integrins αVβ5, CD44, G-protein-coupled receptors (GPCRs), and colony-stimulating factor 1 (CSF1) and its receptor (CSF1R) axis, which collectively regulate macrophage migration (Tang and Guo et al., 2022). Plenty of macrophages have been found in the recurrence of HCC (Liang and Yeh 2014; Zhu and Yang et al., 2019; Tang and Guo et al., 2022), attributed to the stimulation of OPN in within the HCC microenvironment, which subsequently induces macrophage infiltration (Tan and Zhao et al., 2022). SPP1 has been implicated in triggering macrophage polarization towards tumor-associated macrophages (TAMs) with an M2 phenotype, underscoring the complex interplay between OPN, macrophage polarization, and the tumor microenvironment (Liu and Zhang et al., 2022). In addition, programmed death-ligand 1 (PD-L1) was found to be correlated with OPN in hepatocellular carcinoma cells from mice and humans, suggesting OPN plays a role in immune evasion (Wen and Jeong et al., 2016; Song and Chen et al., 2021). OPN will play a partial role in cancer immune surveillance, it will inhibit the proliferation of CD8T cells, inhibit T cell activation, inhibit the cytotoxicity of cytotoxic T lymphocytes, and thus promote tumor growth (Klement and Paschall et al., 2018; Klement and Poschel et al., 2021). Conversely, the findings indicated a carcinostatic or antitumor effect. Fan et al. demonstrated that iOPN in macrophages interacts with the critical Toll-like receptor (TLR) signaling protein MyD88, consequently disrupting TLR signaling pathways and attenuating the synthesis of pro-inflammatory cytokines (Fan and He et al., 2015).
In summary, these findings highlight OPN’s intricate role in HCC progression and its immune environment, suggesting its potential as a therapeutic target for HCC and its metastases. However, additional research is critically needed to substantiate its therapeutic efficacy for HCC.
Alcoholic fatty liver disease
Alcohol-induced damage to the liver remains one of the leading causes of liver disease-related death and transplantation (Chen and Ou et al., 2018; Osna and Rasineni et al., 2022). There is a crucial elevation in OPN levels in the liver and serum of patients diagnosed with alcoholic hepatitis (AH), alcoholic cirrhosis, and end-stage alcoholic fatty liver disease (ALD) (Seth and Gorrell et al., 2006; Morales-Ibanez and Domínguez et al., 2013; Wen and Jeong et al., 2016; Tan and Zhao et al., 2022). A correlation has been identified between serum OPN levels and the severity of AH severity (Morales-Ibanez and Domínguez et al., 2013). Notebly, OPN levels did not change in patients with hepatic alcoholic steatosis, but increased OPN expression has been documented in a rodent model of alcoholic steatohepatitis (ASH) (Banerjee and Burghardt et al., 2006; Wen and Jeong et al., 2016). Banerjee A et al. found that ALD is more prevalent in female rats during ASH, attributed to increased OPN-mediated neutrophil infiltration (Banerjee and Apte et al., 2006). Subsequent research has identified that the N-terminal integrin binding motif (SLAYGLR) of OPN mediates signaling through α4β1 and α9β1 integrins, which may contribute to increased hepatic neutrophil infiltration and activation, as well as exacerbated liver injury in the rat model of ASH (Banerjee and Lee et al., 2008; Wen and Jeong et al., 2016). In addition, several researches suggest that OPN is a protective factor in ASH and AH(Ge and Leunget al., 2014; Lazaro and Wu et al., 2015). SPP1 deficiency does not decrease the incidence of AH or the expression of progenitor and fibrogenic genes; instead, it increases the induction of IL-17 A and polymorphonuclear cell infiltration in certain mice. Moreover, in the absence of SPP1, mice with chronic ASH that do not experience weekly binge episodes begin to develop AH (Lazaro and Wu et al., 2015). The conflicting results may be attributed to the distinct isoforms, iOPN and sOPN (Inoue and Shinohara 2011). Spp1-/- mice lack both isoforms, whereas the application of OPN neutralizing antibodies or aptamers specifically targets and depletes only sOPN (Coombes and Swiderska-Syn et al., 2015). It would be necessary to examine the differences between sOPN and iOPN to determine if OPN is protective in chronic ASH/AH. Interestingly, studies have shown that OPN present in milk can mitigate ethanol-induced liver injury and prevent early alcohol-induced liver damage by blocking lipopolysaccharide translocation in vivo (Lazaro and Wu et al., 2015; Tan and Zhao et al., 2022). In conclusion, further research is warranted to elucidate the complex role of OPN in ALD.
Nonalcoholic fatty liver disease
Nonalcoholic steatohepatitis (NASH) involves fat buildup, liver inflammation, cell damage, and fibrosis, raising the risk of advancing to end-stage liver disease. OPN serves as a biomarker for NASH and fibrosis in individuals with nonalcoholic fatty liver disease (NAFLD) (Wen and Jeong et al., 2016; Glass and Henao et al., 2018). OPN is connected to NAFLD development related to abdominal obesity. In obesity-induced liver steatosis, both OPN expression and serum OPN levels are significantly increased in the liver, which is associated with several metabolic indicators, including the accumulation of hepatic triglycerides (Sahai and Malladi et al., 2004; Nardo and Grün et al., 2020; Yilmaz and Ozturk et al., 2013; Tan and Zhao et al., 2022). In OPN knockout mice, obesity-induced hepatic inflammation and macrophage accumulation were significantly reduced (Tan and Zhao et al., 2022). The absence of OPN appears to mitigate obesity-related hepatic steatosis by promoting insulin receptor substrate-2 (IRS-2) phosphorylation, inhibiting the up-regulation of forkhead box protein O1 (FOXO1) and its tumorigenic target genes, and enhancing hepatic insulin signaling pathway (Bertola and Deveaux et al., 2009; Tan and Zhao et al., 2022). Moreover, normal OPN can reduce obesity-related inflammation in the liver and modulate transcriptional signaling pathways associated with insulin resistance (Kiefer and Neschen et al., 2011; Tan and Zhao et al., 2022). These findings offer novel insights into potential therapeutic targets for the treatment of NASH, indicating that the modulation of OPN or its downstream signaling pathways may represent an innovative therapeutic strategy. However, prior research in the progression of NASH also has yielded inconsistent findings. Macrophage-derived osteopontin provides protection against NASH by upregulating oncostatin M (OSM). This upregulation subsequently enhances the expression of arginase 2 (ARG2) via the signal transducer and activator of transcription 3 (STAT3) signaling pathway. The increase in ARG2 activity facilitates enhanced fatty acid oxidation, thereby contributing to a reduction in hepatic steatosis (Han and Ge et al., 2023).
Pancreatic diseasesPancreatic cancer
OPN is markedly over-expressed in pancreatic cancer and plays a critical role in promoting the proliferation, migration, and invasion of cancer cells. Its interaction with integrins and CD44 on cell surface is essential for facilitating the adhesive, survival, proliferative, and metastatic capabilities of these cells (Lu and Liu et al., 2021). Notably, nicotine, a principal component of cigarette smoke and a significant risk factor for pancreatic cancer, has been shown to induce the expression of OPN. This finding suggests that OPN may serve as a direct link between smoking and the progression of pancreatic cancer (Chipitsyna and Gong et al., 2009). Furthermore, the interplay between OPN and the Hedgehog signaling pathway has been identified as a promoter of pancreatic tumor growth and a mechanism conferring resistance to apoptosis (Wu and Yang et al., 2021). Subsequent research has demonstrated that the SPP1 are significantly enriched with H3K4me3 in the pancreatic cancer genome, facilitating immune evasion and resistance to anti-PD-1 therapy (Lu and Liu et al., 2021). Targeting OPN in pancreatic cancer therapy may hold significant clinical value, with potential strategies involving OPN-specific antibodies or small molecule inhibitors to impede pancreatic tumor progression and metastasis. The integration of immune checkpoint inhibitors, such as PD-L1, with OPN-targeted therapies may represent an innovative therapeutic strategy.
Chronic pancreatitis
In chronic pancreatitis, OPN plays a pivotal role in the activation of pancreatic stellate cells (PSCs) and the ensuing fibrotic development. OPN engagement with integrin receptors on PSCs fosters their activation and enhances the synthesis and secretion of the extracellular matrix (Cao and Liet al., 2019). Additionally, OPN is implicated in modulating inflammatory responses in chronic pancreatitis by promoting the recruitment and activation of inflammatory cells, including macrophages and neutrophils. These cells release substantial quantities of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6, thereby exacerbating pancreatic inflammation (Sarosiek and Jones et al., 2015).
Pancreatic islet function
In the context of diabetes, particularly in models induced by streptozotocin, OPN expression is up-regulated, highlighting its potential role in islet cell protection (Katakam and Chipitsyna et al., 2005). The supplementation of OPN from external sources notably augmented the insulin release in response to glucose in individuals with diabetes, contrasting with those maintaining normal blood sugar levels. Dickerson MT et al. indicate that the TALK-1/iOPN complex induces membrane hyperpolarization and diminishes glucose-stimulated calcium influx in β-cells, which is anticipated to impede glucose-stimulated insulin secretion (Dickerson and Vierraet al., 2017). These findings offer corroborating evidence for the protective function of OPN on islets under diabetic conditions, and may identify novel therapeutic targets for islet preservation in Type 2 Diabetes.
Biliary diseasesCholangiocarcinoma (CCA)
Increased expression of OPN in CCA is correlated with heightened tumor aggressiveness, metastatic capacity, and unfavorable prognosis for patients (Laohaviroj and Chamgramol et al., 2016). At the molecular level, OPN interacts with mitogen-activated protein kinase 1 (MAPK1) and facilitates the phosphorylation of β-catenin at Ser675, thereby activating the Wnt signaling pathway, which promotes tumor cell proliferation and invasion (Zheng and Zhou et al., 2018). OPN’s interplay with the tumor microenvironment is also significant, as it engages with cancer-associated fibroblasts and facilitates the progression of CCA through the NR4A2/OPN/Wnt signaling axis (Jing and Fu et al., 2021). OPN may promote angiogenesis in CCA by modulating the expression of vascular endothelial growth factor (VEGF), aiding tumor growth and dissemination (Cadamuro and Sarcognato et al., 2023). Furthermore, Zhou KQ et al. show that low circulating OPN levels might signal aggressive traits and poor prognosis in ICC patients, affecting chemotherapy effectiveness. High OPN expression could hinder ICC metastasis by downregulating MMP1, MMP10, and CXCR4. Thus, circulating OPN levels could serve as a biomarker for early CCA detection, disease monitoring, and assessing therapeutic response (Zhou and Liu et al., 2019).
Primary sclerosing cholangitis (PSC)
In Primary Sclerosing Cholangitis (PSC), a chronic inflammatory condition affecting the bile ducts, OPN has been identified a marker for bile duct-associated macrophages and demonstrates a correlation with the severity of liver fibrosis (Yang and Ramachandran et al., 2014). Especially in the co-existence model of cholangitis and colitis, the expression of OPN is intimately connected with the alterations in hepatic macrophages. OPN has the potential to be a predictive biomarker for survival without liver transplantation in PSC patients. This is supported by the heightened serum levels of OPN in PSC patients compared to the control group, with a significant surge observed in the advanced stages of the disease (De Muynck and Heyerick et al., 2024).
Biliary atresia and obstructive cholestasis
OPN has been implicated as a potential biomarker for assessing the severity and progression of biliary atresia (Whitington and Malladi et al., 2005). In cases of obstructive cholestasis, caused by bile duct obstruction, OPN acts as an early mediator of inflammation and hepatic injury. Its expression is markedly increased following bile duct ligation in murine models, associated with liver damage and neutrophil recruitment. Additionally, matrix metalloproteinases (MMPs) activate OPN, generating pro-inflammatory forms that exacerbate inflammatory responses and hepatic injury (Yang and Ramachandran et al., 2014). These findings indicate that OPN plays a role in the signaling pathways initiated by bile duct epithelial cells and may have a regulatory effect on inflammation associated with obstructive cholestasis.




Comments (0)