
Remember me


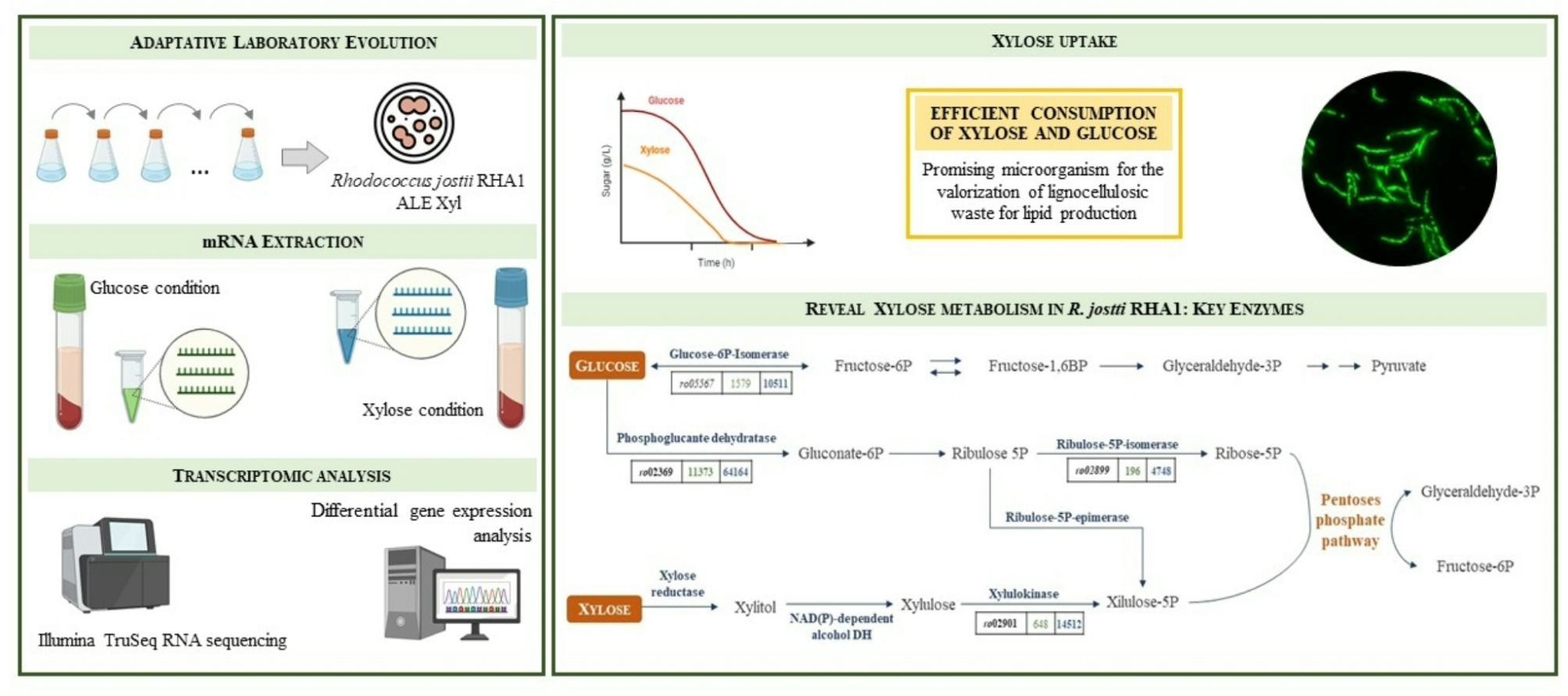



Based on the assumption that enzyme promiscuity and phenotypic flexibility of microorganisms shape adaptation to novel growth substrates [32], and the fact that R. jostii RHA1 natively has two xylB genes that can facilitate the consumption of xylose [14], we developed an ALE strategy to test if RHA1 can be evolved to efficiently metabolize xylose without expressing heterologous enzymes.
When we cultured R. jostii RHA1 in minimal media containing xylose as the sole carbon and energy source we observed a slight growth after 1 week of cultivation. After four passes, we were able to isolate one clonal strain named ALE-xyl able to grow very efficiently on minimal medium agar plates containing xylose. Figure S4 shows the growth of the wild type RHA1 and ALE-xyl strains in rich medium as well as in minimal media containing xylose as sole carbon and energy source. The identity of the ALE-xyl strain as a derivative of RHA1 was confirmed by its 16 S RNA sequence (data not shown).
To determine the stability of the xylose phenotype of the ALE-xyl strain, the isolated clone was cultured in rich medium and then transferred again to minimal medium containing xylose as sole carbon source. Figure S5 shows that ALE-xyl strain progressively loses the ability to grow on xylose when cultured in rich medium. The strain only maintains the ability to grow on xylose when cultured in minimal medium containing xylose suggesting that the capacity of ALE-xyl strain to metabolize this monosaccharide is most probably due to a reversible phenotypic adaptation.
Xylose consumption by ALE-xyl strainThe growth on xylose of ALE-xyl strain was compared with that of the recombinant strain R. jostii RHA1 (pNVSxylABatf1), harbouring the xylose isomerase pathway, which was constructed in our lab to overproduce lipids using xylose as substrate. Figure 2 shows that both strains are able to grow when cultured in minimal W medium containing 30 g/L of xylose as the sole carbon and energy source. However, the xylose consumption by the ALE-xyl strain was higher than that observed in the recombinant strain which was not evolved by ALE.
Fig. 2
Time course of R. jostii RHA1 ALE-xyl and R. jostii RHA1 (pNVSxylABatf1) strains cultured in xylose. (A) Xylose consumption. (B) pH. (C) Optical density. (D) Total sugar consumption. The culture was carried out with xylose (30 g/L)
We observed that when cells are cultured in flasks on xylose the pH of the medium decreases as far as xylose is consumed and when pH drops below 5 the strains stop growing (Fig. 2B), indicating that a strict control pH is critical to facilitate the complete consumption of xylose.
Figure 3 shows how ALE-xyl strain produces more lipids (54%) than the recombinant strain (19%) according to the percentage of FAMEs in relation to the total biomass. Moreover, FAMEs profile produced by both strains is very similar, being palmitic acid the most abundant fatty acid in both cases. Figure S6 shows microscopy images of lipids accumulated and produced by both strains.
Fig. 3
(A) Lipid content of R. jostii RHA1 ALE-xyl and R. jostii RHA1 (pNVSxylABatf1) strains cultured on media with xylose as sole carbon source. (B) FAMEs of ALE-xyl and R. jostii RHA1 (pNVSxylABatf1) strains cultured on media with xylose as sole carbon source
Growth of the ALE-xyl strain in a lignocellulosic-like hydrolysateTo determine if ALE-xyl strain could be able to consume at the same time glucose and xylose in a lignocellulosic-like hydrolysate (glucose + xylose + arabinose), the strain was cultured in minimal W medium containing a synthetic mixture of glucose, xylose and arabinose mimicking the proportion that can be found in a saccharified extract of a corn stove prepared in our laboratory (glucose (27 g/L), xylose (7 g/L) and arabinose (1.2 g/L). Figure 4 shows how ALE-xyl is able to metabolize xylose in the presence of glucose. It is very interesting that in this evolved strain glucose does not repress the consumption of xylose. Moreover, as expected, arabinose is not metabolized by ALE-xyl, but interestingly, we have observed that it does not interfere with the consumption of glucose or xylose. The lipid production in the ALE-xyl strain in this medium was slightly lower (60%) than the percentage achieved when the strain was cultured using xylose as sole carbon and energy source.
Fig. 4
Time course of R. jostii RHA1 ALE-xyl and R. jostii RHA1 (pNVSxylABatf1) cells cultured in a mixture of sugars (glucose (20 g/L), xylose (6 g/L) and arabinose (1 g/L).): (A) Optical density at 600 nm; (B) pH; (C) glucose consumption; (D) xylose consumption; (E) arabinose consumption
Transcriptomic analysisOnce we had demonstrated that the ALE-xyl strain was capable of growing efficiently and producing lipids using xylose as the sole source of carbon and energy, we set out to investigate which genes could be responsible for the phenotypic adaptation that occurred in this strain. To this aim, we performed a comparative transcriptomic analysis of the ALE-xyl strain cultured in minimal medium containing glucose or xylose as a sole carbon and energy source (see supplementary material). These analyses revealed that xylose growing cells highly overexpressed the putative operon RHA1_02898 – RHA1_ro02909 that we have named as pen operon (Fig. 5; Table 1). The function of this operon has not been previously described in Rhodococcus or in other microorganisms, but the annotated functionality that can be ascribed to the genes encoded in this operon suggests that it can be involved in the metabolism of pentose sugars. A systematic search on the genome data bases revealed that the same operon is present in several Rhodococci and in other closely related actinomycetes (Fig. 5).
Fig. 5
Scheme of the pen operon (RHA1_ro02898 - RHA1_ro02909) overexpressed in R. jostii RHA1 ALE-xyl strain growing in xylose. This operon is also present in microorganisms of Actinobacteria phylum
Table 1 Genes from the Pen Operon (RHA1_ro02898 - RHA1_ro02909) overexpressed in R. jostii RHA1 ALE-xyl strain growing in xyloseAlthough the RHA1_ro02897 gene encoding a putative acyl-CoA thioesterase located at the 3’-end of the pen operon is also overexpressed (Table 1), this gene does not seem to form part of the pen operon. The distance between the stop codon of the last gene of the operon and the start codon of the thioesterase encoding gene is too long (i.e., 194 bp), suggesting that this gene is expressed by an independent promoter (Fig. 5). In contrast, the distance within the stop and start codons of the contiguous genes contained in the pen operon ranges between 1 and 55 bp. The reason why this thioesterase is overexpressed, even when it is not apparently related to sugar metabolism, is unknown, nevertheless, we have detected a long palindromic region (GTTGTAGCGTACAAC) overlapping the boxes − 35 and − 10 of the gene (Fig. 5) that could be recognized as a consensus operator sequence by the PenR regulator.
Remarkably, the pen operon contains the RHA1_ro02901 (penH) gene encoding the xylulokinase described by Xiong et al. (2012) to be functional on xylulose. However, the other xylulokinase (RHA1_ro02812 gene) annotated in the genome of R. jostii RHA1 and also described by Xiong et al. (2012) as functional on xylulose was apparently expressed constitutively [14]. This finding can explain why RHA1_ro02812 is more efficient in the wild type strain than RHA1_ro02901 [14], because the last remain silent.
Remarkably, the pen operon encodes an ABC transporter (penABC genes) that is homologous to pentose transporters and that might facilitate the uptake of xylose. Although there are other annotated sugar transporters in the genome of RHA1, none of them are overexpressed in the presence of xylose. Even more the RHA1_ro02804-RHA1_ro02808 genes encoding the only sugar ABC transporter that is overexpressed in glucose show expression levels far below those observed for the penABC genes (Table 2). Nevertheless, we cannot discard that other less expressed putative sugar transporters can contribute to the uptake of xylose.
Table 2 Sugars transporters expressed in R. jostii RHA1 ALE-xyl strain under glucose and xylose growth conditionsA FadR/GntR-like regulator, named as PenR, that is strongly expressed in the presence of xylose is located just upstream of the operon suggesting that the expression of the pen operon can be controlled by this regulator. An analysis of the promoter region suggests that most probably the mRNA of PenR does not have a Shine-Dalgarno sequence, because there is not a typical consensus sequence preceding its start codon and because the − 10 box of the putative promoter is located very close to the start codon. Therefore, this promoter could render a typical leaderless transcript frequently found in actinomycetes [33].
The in-silico analysis of the promoter region of the pen operon revealed that within the putative − 10 box there is a perfect palindromic GTATAC sequence that matches with the palindromic NyGTNxACNy consensus sequence proposed as operator for the GntR-like regulators [34] (Fig. 5). According to this observation we could propose that the PenR regulator might act as a repressor. Nevertheless, considering that PenR is overexpressed in ALE-xyl we cannot discard that this regulator can be an activator since GntR-like regulators can act either as repressors or activators.
Although the genome of R. jostii RHA1 contains many genes annotated as sugar isomerases none of these genes appear to be overexpressed in xylose and thus, we cannot propose that an isomerase could be responsible for metabolizing xylose through the typical isomerase pathway frequently found in bacteria. However, we cannot discard that the basal expression of one of these isomerases, that in general accept a wide range of substrates, can be enough to transform xylose into xylulose once the sugar is uptake by the ABC pentose transport system contained in the operon.
Figure 6 shows the differential expression of the annotated genes involved in the metabolism of hexoses and pentoses in R. jostii RHA1 when the ALE-xyl cells are cultured in glucose or xylose. These results strongly suggest that xylose might be metabolized in RHA1 strain through a reductase pathway as proposed for Shewanella oneidensis [35]. Interestingly, RHA1 contains at least three annotated putative aldolase/keto reductases encoded by the genes RHA1_ro04589, RHA1_ro05024, and RHA1_ro00567 that can act as xylose reductases, and in fact some of them are constrictively and highly expressed in ALE-xyl, both in glucose and xylose containing media (See counts at Fig. 6). We have also observed a putative alcohol dehydrogenase encoded by the gene RHA1_ro02809 that is highly expressed in glucose and xylose that might act as a xylitol/xylulose dehydrogenase (Fig. 6). Nevertheless, we cannot discard that other alcohol dehydrogenases could play this role as suggested by Sekar et al. (2016) [35].
Fig. 6
Expression levels of the genes involved in the first steps of the metabolism of glucose and xylose in R. jostii RHA1. The data are presented in counts
Figure 6 shows that the glucose-6-P isomerase and the glucose-6-P reductase are overexpressed in the presence of xylose suggesting that the ribose-5-P required for the pentose pathway can be produced from the isomerization of ribulose-5-P generated by the oxidative reactions of the pentose pathway. A putative ribos-5-phosphate isomerase (RHA1_ro02899 gene, rpiA-like) is encoded in the pen operon, reinforcing this hypothesis. Thus, we propose that half of the fructose-6-P produced by the non-oxidative reactions of the pentose pathway can be transformed by glucose-6-P isomerase into glucose-6-P that is further transformed by the glucose-6-P reductase into 6-phosphogluconolactone. Nevertheless, we cannot completely discard that ribose-5-P can be produced from xylulose-5-P through the serial action of ribulose-5-P epimerase that generates ribulose-5-P and the ribose-5-P isomerase that transforms the ribulose-5-P into ribose-5-P.
The transcriptomic analysis also revealed that the Entner-Doudoroff pathway is also functional in ALE-xyl either in glucose or xylose (Table S1). In R. jostii RHA1 the genes responsible for this pathway are organized in three operons within the RHA1_ro02362- RHA1_ro02370 cluster. Interestingly, the RHA1_ro02365 gene encoding a sugar MFS facilitator has been described as the main glucose transport system in RHA1 [36]. Considering that these genes are highly expressed in the presence of xylose we must assume that the regulation of these genes should be carried out by glucose-6-P or any other intermediate of the pathway. Interestingly, this cluster does not encode a gluconolactonase-6-P activity that is located elsewhere in the genome (RHA1_ro07182), but low expressed (Fig. 6), suggesting that gluconolactone-6-P can be also spontaneously transformed into gluconate-6-P as suggested by other authors [37].
Considering all alternative pathways, several types of metabolic reactions can take place in RHA1:
$$\begin3}Xylose\, \to \,G3P\, + \,2\,F6P \hfill \\\,\,\,\,\,\,\,\,\,\,\,\,}\left( }epimerization - isomerization} \right) \hfill \\ \end $$
$$\begin2}Xylose \to \,C\, + \,G3P\, + \,F6P\, \hfill \\\,\,\,\,\,\,\,\,\,\,\,\,\,\left( }oxidative}pentose}reactions} \right) \hfill \\ \end $$
F6P → G3P and pyruvate (Entner-Doudoroff or Embden-Meyerhoff (glycolysis) pathways).
Surprisingly, we have observed that in the presence of xylose many of the S30 and S50 ribosomal genes are overexpressed when compared with the expression levels observed in glucose, suggesting that the metabolism of xylose requires the synthesis of a larger number of ribosomes (see Table S2). In this sense, ribosome abundance has been correlated with growth rate in C. glutamicum [38].
Finally, it is worth to mention that the RHA1_ro04085-RHA1_ro04093 gene cluster that is annotated as a rhamnose metabolic cluster is not expressed neither in glucose or xylose, strongly suggesting that these genes are not involved in the composition of xylose by the ALE-xyl strain.
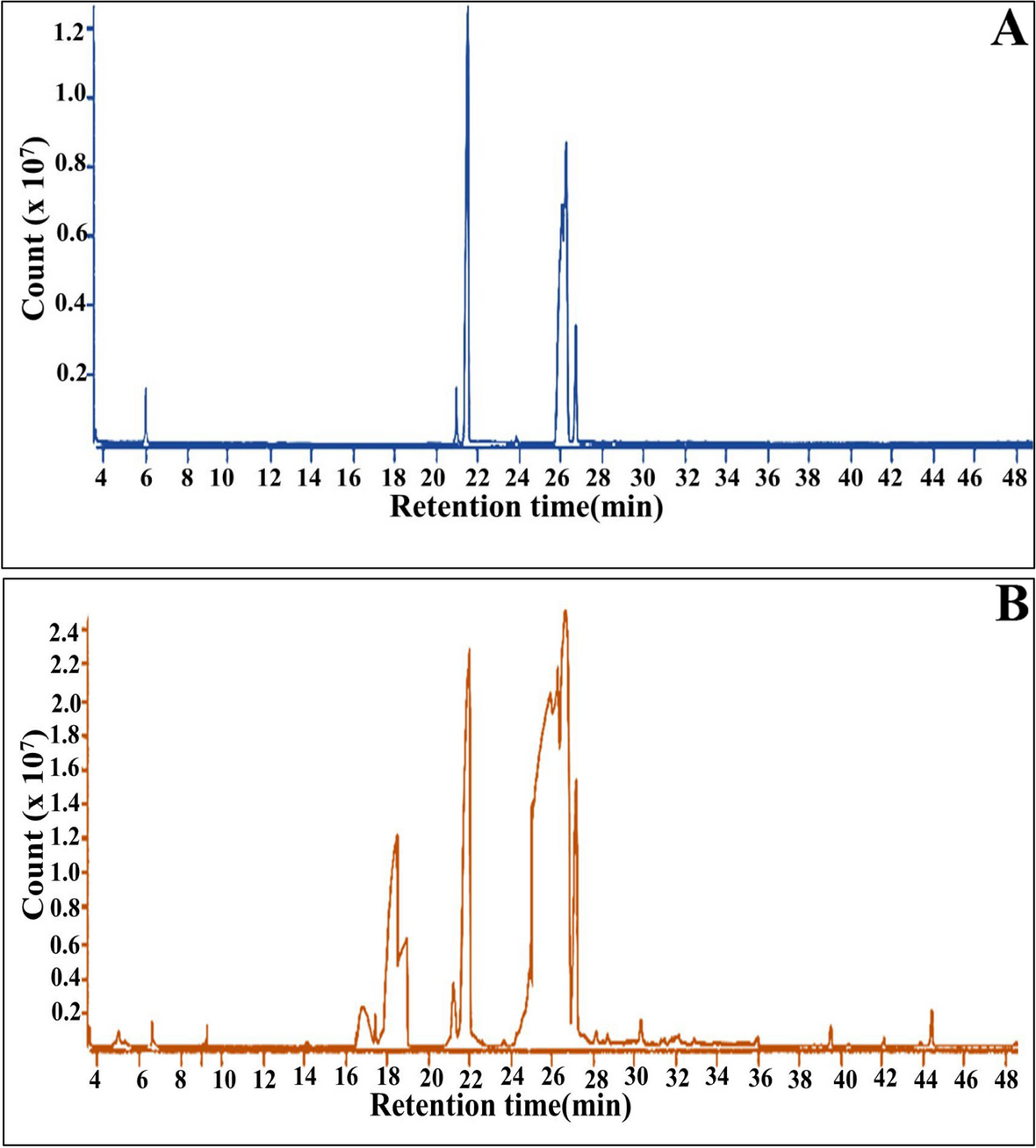
Assay of xylose reductase activityTo determine if the ALE-xyl cells growing in xylose express a xylose reductase, a crude extract of these cells was analysed to determine the presence of this activity. Figure S7 shows that xylose reductase activity is detectable in the crude extract, indicating that xylose can be converted into xylitol. Figure S7 also shows that as usual xylose reductase can utilize both NADH and NADPH as cofactors.
Interestingly, when the ALE-xyl crude extract was analysed by HPLC to detect the presence of xylitol we observed a pick in the chromatogram corresponding to a small amount xylitol (0.5 mg/mL of extract), which agrees with the expression of the xylitol reductase.
Comments (0)