
Remember me

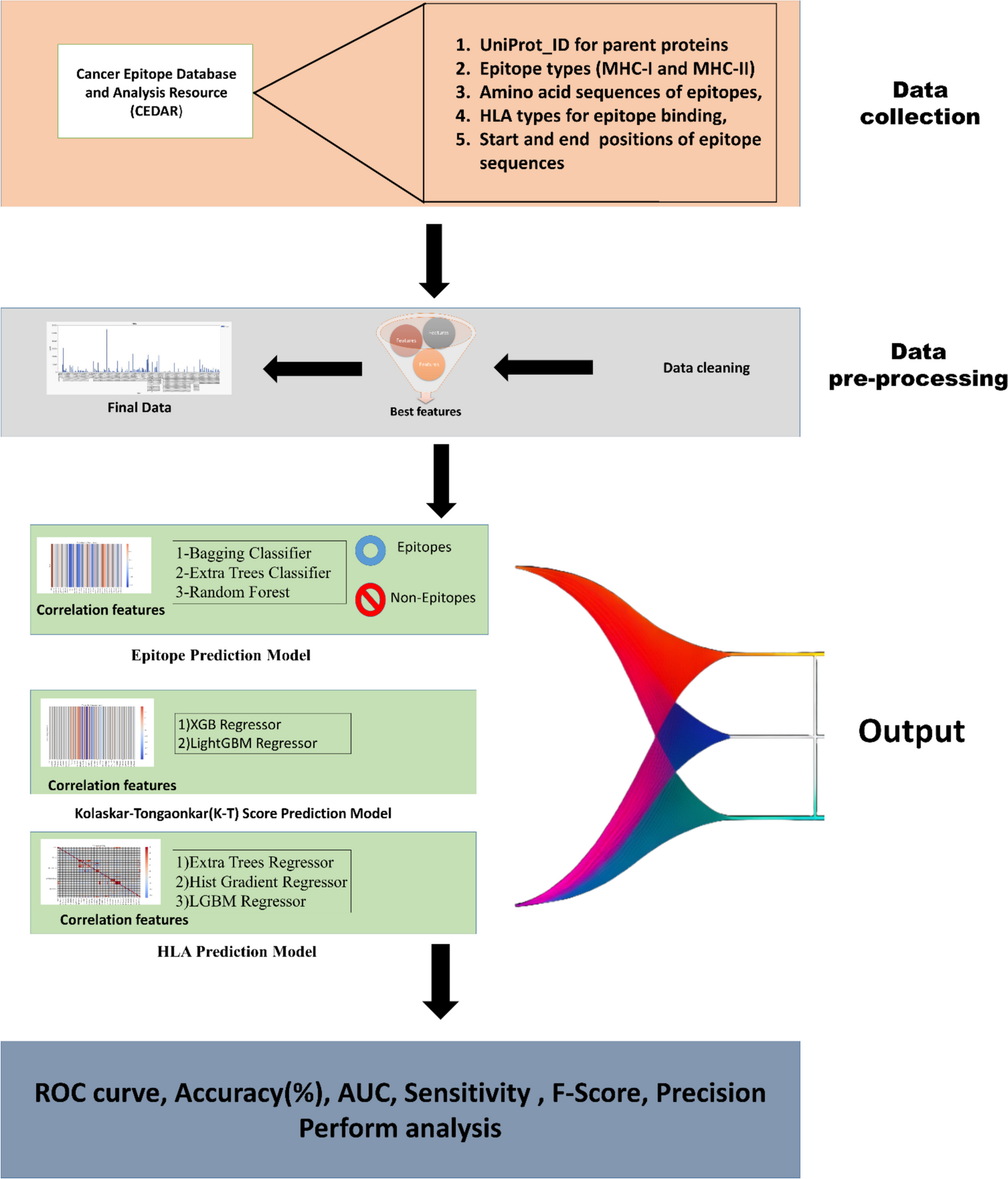





Investigation of the TLR9 expression across HEL, SET2, and K562 cell lines revealed dynamic response when treated with ON-GNPs targeting wild-type JAK2 (JAK2), JAK2V617F, and scramble control. The relative expression of TLR9 was assessed at short-term (0.5–2 h) and long-term (24–72 h) time points, as shown in Fig. 1A, B, and C. Overall, TLR9 expression remained stable across all treatments during the first 24 h. However, a significant increase in TLR9 expression was observed between 24 and 48 h, which then returned to baseline levels by 72 h of incubation.
Fig. 1
Relative expressional change of TLR9, IRF7, and NFKB1 genes upon varying types of GNP application (complementary to JAK2V617F mRNA-specific, JAK2 mRNA-specific, and scrambled-non-specific to any mRNA) in cell lines of JAK2V617F biallelic HEL (A, D, G), JAK2V617F monoallelic SET2 (B, E, H), and JAK2V617F-negative K562 (C, F, I) for short-term (0.5–2 h) and long-term (24–72 h) durations (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001)
The most pronounced change in TLR9 expression was detected in SET2 cells, where a substantial increase (~ 200–300-fold, p < 0.0001) occurred at 24 and 48 h before sharply declining at 72 h (Fig. 1B).
This suggests a cell type-specific response mechanism in which the megakaryoblastic lineage of SET2 cells may enhance their sensitivity to ON-GNP treatment in the long term. Differences in transcriptional regulation, nucleic acid-sensing pathway activity, and cytokine feedback loops may contribute to this heightened response. In contrast, HEL cells (erythroblastic) and K562 cells may rely on distinct regulatory mechanisms that modulate inflammatory signaling differently. Interestingly, no significant differences in relative TLR9 expression were observed between treatments with ON-GNPs targeting JAK2, JAK2V617F, or the scramble control, indicating that the observed effects are likely not dependent on the specific target of the ON-GNPs.
A parallel assessment of relative IRF7 expression under similar experimental conditions (Fig. 1D, E, and F) revealed minimal changes during the early phase (0.5–2 h) of exposure. However, a significant increase in IRF7 expression was detected at 24 and 48 h and subsequently normalized by 72 h. Notably, JAK2V617F-GNPs induced a markedly higher expression of IRF7 in HEL and SET2 cells during these intervals with fold changes of approximately 20 (p < 0.001) and 100 (p < 0.0001), respectively. This pronounced response highlights the potential influence of genetic variations, such as the presence of JAK2V617F, on cellular responses to ON-GNP treatment (Fig. 1D, E).
The marked upregulation of IRF7, particularly in JAK2V617F-positive HEL and SET2 cells, highlights the critical role of type I interferon responses in nucleic acid sensing within the context of MPNs. This finding aligns with previous reports of heightened interferon signaling associated with JAK2V617F-driven inflammation. Overall, the transient activation of TLR9 and IRF7, followed by their normalization, suggests that ON-GNPs may recalibrate dysregulated inflammatory pathways in MPNs, reducing chronic inflammation without prolonged immune suppression.
Investigations on NFKB1 expression revealed interesting results. In all cell lines, short-term exposure (0.5–2 h) to ON-GNPs had no effect; however, a notable increase was observed at 24 h across all ON-GNP treatments, which diminished by 72 h. HEL and SET2 cells exhibited an earlier but transient increase in NFKB1 expression significantly elevated at both 24 h and 48 h (~ tenfold) in response to JAK2- and JAK2V617F-GNPs before declining (Fig. 1G, H). Interestingly, K562 cells displayed a delayed and selective response, with NFKB1 upregulation occurring only at 48 h, showing a ~ 30-fold increase (p < 0.001) with JAK2-GNP and a ~ 20-fold increase (p < 0.001) with scramble-GNP (Fig. 1I). The significant increase in NFKB1 expression following JAK2-GNP treatment suggests a potential interplay between JAK2-targeting ON-GNPs and BCR-ABL-driven pathways. This effect may arise from off-target interactions, secondary inflammatory signaling activation, or indirect modulation of NF-κB via altered cytokine dynamics.
Delayed activation of cGAS-STING pathway with complementary JAK2 and JAK2V617F mRNAs attached gold nanoparticlesTo investigate the role of alternative nucleic acid-sensing pathways in intracytoplasmic activities, we analyzed the expression of key cGAS/STING pathway genes, including cGAS, STING, TBK1, and IRF3, in response to ON-GNP treatment.
Prolonged exposure (72 h) to ON-GNPs significantly increased cGAS expression across all three cell lines as shown in Fig. 2A–C. HEL cells showed a marked upregulation of cGAS with both JAK2-GNP (p < 0.001) and JAK2V617F-GNP (p < 0.0001) (Fig. 2A). In SET2 cells, a significant increase was observed only with scramble-GNP at 24 h and 48 h (p < 0.0001) and JAK2-GNP at 72 h, while JAK2V617F-GNP elicited no substantial effect (Fig. 2B). In K562 cells, relative cGAS expression profoundly increased (~ 40-fold, p < 0.01) at 2 h and 48 h with JAK2V617F-GNP (p < 0.001) and at 72 h with JAK2-GNP (p < 0.0001) (Fig. 2C). These findings suggest a cell line- and treatment-specific regulation of cGAS expression. However, STING expression remained mostly unaffected by ON-GNP treatment, with isolated exceptions. In HEL cells, STING was significantly upregulated at 48 h with JAK2V617F-GNP (p < 0.05) (Fig. 2D), and in SET2 cells, STING expression increased at 1.5 h with the JAK2V617F-GNP (p < 0.05) and at 24 h (p < 0.01) and 48 h (p < 0.001) with scramble-GNP (Fig. 2E). In K562 cells, no substantial STING response was detected despite strong cGAS induction (Fig. 2F). These results indicate that in SET2 cells, STING activation occurs only in response to non-specific ON-GNP treatment (scramble-GNP), whereas JAK2- and JAK2V617F-targeting ON-GNPs had no effect on STING expression.
Fig. 2
Relative expressional change of cGAS (A, B, C), STING (D, E, F), TBK1 (G, H, I), IRF3 (J, K, L), and JAK2 (M, N, O) in cell lines HEL, SET2, and K562 upon incubation with JAK2, JAK2V617F, and scramble-GNPs for short-term (0.5–2 h) and long-term (24–72 h) durations. (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001)
TBK1 expression was generally stable, with minor exceptions, such as increased expression in SET2 at 2 h with JAK2V617F-GNP (p < 0.001) and in K562 at 2 h with JAK2V617F-GNP (p < 0.001) (Fig. 2G–I). IRF3 expression, in contrast, demonstrated early stage sensitivity to ON-GNP treatment in HEL and SET2 cells at 2 h with JAK2V617F-GNP (~ 13-fold, p < 0.05) and at 1.5 h with JAK2-GNP (~ 30-fold, p < 0.0001), respectively. HEL cells exhibited a delayed but significant response at 48 h with JAK2V617F-GNP (p < 0.05) (Fig. 2J, K). In contrast, K562 cells exhibited early stage IRF3 upregulation with JAK2-GNP treatment, starting at 0.5 h and persisting until 2 h, before diminishing at 24 h until 72 h. JAK2-GNP treatment induced IRF3 after (Fig. 2L).
The dynamics of JAK2 expression following ON-GNP treatment underscored the complex regulatory mechanisms. Short-term suppression was followed by significant upregulation at 72 h in HEL cells treated with JAK2V617F-GNP (p < 0.01) indicating a potential compensatory mechanism in response to targeted suppression (Fig. 2M). In SET2 cells, JAK2 expression significantly increased with JAK2-GNP at 24, 48, and 72 h (p < 0.0001, p < 0.001, and p < 0.0001, respectively) (Fig. 2N). Surprisingly, significant changes were observed in K562 cells (Fig. 2O), underscoring the specificity of ON-GNPs to JAK2 contexts with early induction within 0.5–2 h following JAK2V617F-GNP and scramble-induced activation in 1.5–2 h. The JAK2 activation diminished by 24 h and lasted until 72 h. Further studies are needed to elucidate whether ON-GNP treatment influences additional regulatory elements within the BCR-ABL and JAK2 signaling pathways.
This analysis demonstrates the intricate cellular responses to ON-GNPs and highlights the significant influence of genetic background and cell origin on gene expression dynamics. These findings suggest that ON-GNP-mediated interventions have the potential to modulate nucleic acid-sensing pathways and gene expression in a cell- and mutation-specific manner, providing insights into their utility as targeted therapeutic strategies in the context of MPNs.
The effect of complementary JAK2V617F mRNAs attached gold nanoparticle stimulation on TLR9 and RAGE receptorsTo investigate the mechanisms underlying the transient suppression and subsequent upregulation of TLR9 expression in response to ON-GNPs targeting the complementary transcripts of JAK2V617F, we analyzed the role of nucleic acid sensors, specifically TLR9 and RAGE. Nucleic acids and RAGE interact at the cell surface, promoting their uptake via the endosomal route, where the TLR9 receptors are predominantly located. The study aimed to distinguish the activation patterns of TLR9 and RAGE, hypothesizing that ON-GNPs may influence the TLR9/RAGE pathway and modulate the expression of other genes in a stimulus-duration-dependent manner.
Flow cytometry analysis of HEL cells incubated with JAK2 and JAK2V617F-GNPs revealed no significant changes in intracytoplasmic TLR9 expression at either 2 h (short term) or 24 h (long term) of stimulation (Fig. 3A). Similarly, cell surface and intracytoplasmic RAGE protein levels remained unchanged after both short-term and long-term incubation with JAK2V617F-GNPs (Fig. 3B).
Fig. 3
A Intracytoplasmic staining of TLR9 protein in HEL cells upon 2 h and 24 h incubation with JAK2V617F-GNPs and JAK2-GNPS by flow cytometer. B Intracytoplasmic and cell surface staining of RAGE protein for 2 h and 24 h incubation with JAK2V617F-GNP
These findings highlight the complexity of TLR9 regulation and suggest that TLR9 expression observed in response to ON-GNPs may not directly involve RAGE-mediated pathway, indicating alternative mechanisms of regulation.
The effect of complementary JAK2 and JAK2V617F mRNAs attached gold nanoparticle stimulation on inflammatory cytokinesTo evaluate the impact of ON-GNPs on cytokine secretion in HEL and SET2 cells over extended periods, specifically at 24, 48, and 72 h, we employed a method that facilitated the quantification of various cytokines, including IL-1β, IFN-α2, IFN-γ, TNF-α, MCP-1, IL-6, IL-8, IL-10, IL-12p70, IL-17A, IL-18, IL-23, and IL-33, as shown in Fig. 4. Notably, a significant elevation in IL-8 levels was recorded in the HEL culture medium across all tested conditions, with the peak concentration observed at the 48-h mark following exposure to JAK2-GNPs. The cytokine profiles of SET2, presented in the figure, further complemented these findings.
Fig. 4
Inflammatory cytokine concentration levels of HEL and SET2 cells that were treated with JAK2 mRNA-specific, JAK2V617F mRNA-specific, and scrambled (non-specific to any mRNA)-GNPs detected with inflammation panel
Comments (0)