
Remember me




To determine the effects, if any, of IMR on inflammation and the development of dermatitis, female mice were continuously maintained on one of two high-fat diet regimens, these being either (1) continuous feeding with a methionine-replete control diet (control-fed; CF) or (2) an IMR regimen characterized by alternating feeding of the CF diet (4 days) and a diet containing no sulfur amino acids (3 days) (Supplemental Table 1). Both regimens were initiated when animals were 12 months of age and continued until they displayed evidence of dermatitis. The experiment continued until all animals developed dermatitis, at which point high-fat diet feeding was discontinued, allowing for resolution of the dermatitic lesions. For all cases of dermatitis, frank pruritic lesions were observed on the skin of the dorsal neck, shoulders, and ears. Scratching was a clear contributor to the severity of these lesions, resulting in alopecia, ulcerations, and serocellular crusts (Fig. 1A and Fig. S1). The observed skin condition was thus consistent with murine idiopathic ulcerative dermatitis, which is commonly observed for C57BL/6J mice and related strains and known to be exacerbated by consumption of a high-fat diet [10,11,12]. The median duration of dermatitis-free survival for control-fed animals was 341 days (Fig. 1B). In contrast, mice undergoing IMR demonstrated a significant delay in the development of dermatitis, with a 151 day increase in median dermatitis-free survival (492 days vs. 341 days; p = 0.0449).
Fig. 1
IMR protects high-fat diet-fed mice against dermatitis and chronic inflammation. Shown are (A) representative images of mice that were either control-fed (CF) or intermittently methionine-restricted (IMR), as well as (B) the longitudinal incidence of dermatitis for such animals. The average plasma levels of the inflammation markers (C) haptoglobin and (D) serum amyloid A (SAA) in animals fed the indicated diets are also provided, as well as those of the pro-inflammatory cytokine (E) interleukin-6 (IL-6). As a further assessment of systemic inflammation in these animals, white blood cell (WBC) concentrations were also determined (F). For panels B-E, N = 4 for each group; for panel F, N = 6 for each group. For panel B, the difference between treatment groups is statistically significant (p = 0.0355). For panels C-F, bars denote standard error of the mean (SEM) and statistically significant differences are indicated (**, p < 0.01; ****, p < 0.0001)
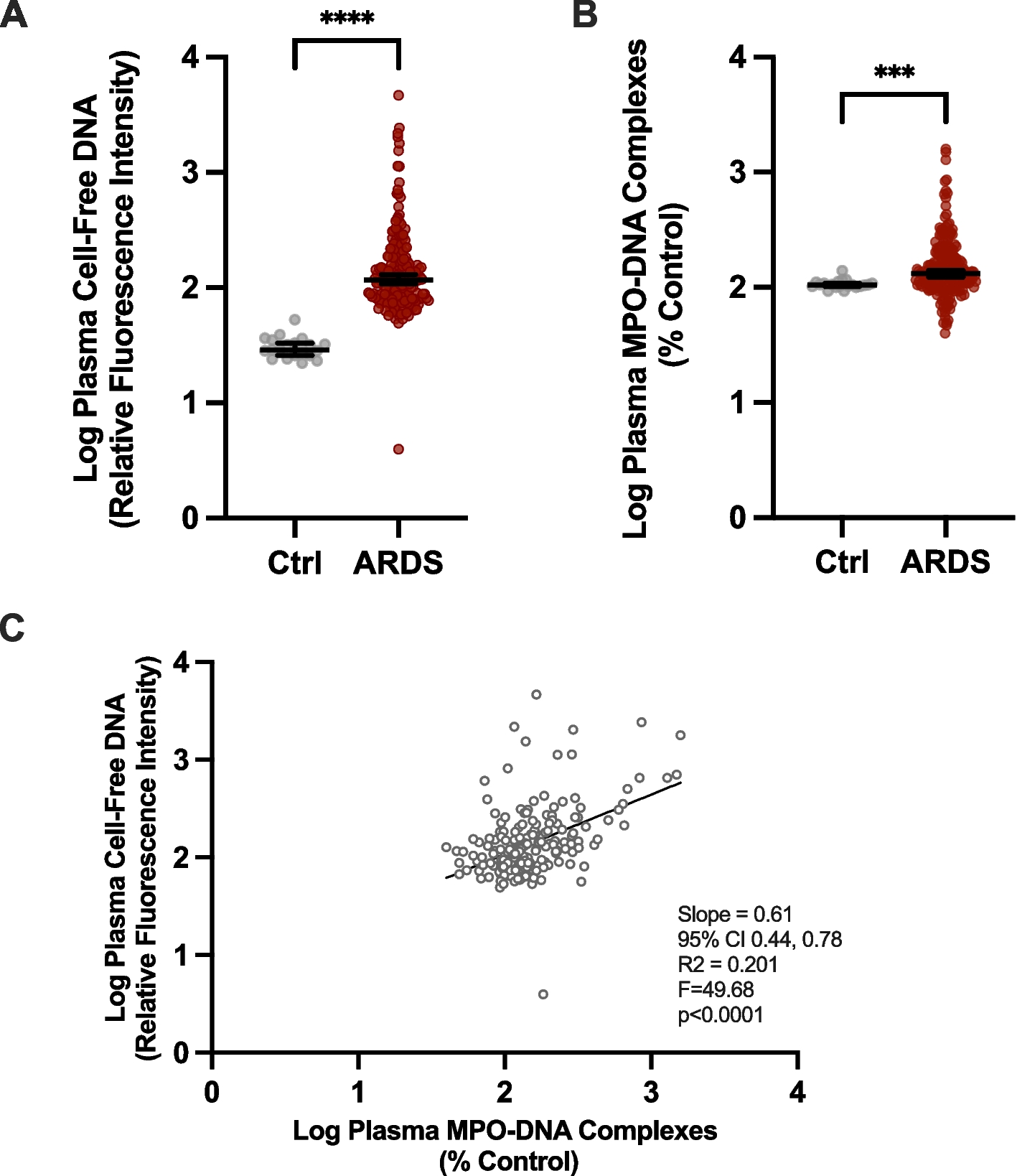
To determine whether, as hypothesized, the observed protection against dermatitis conferred by IMR was characterized by reduced systemic inflammation, we assessed the circulating levels of two acute phase reactants, haptoglobin and serum amyloid A (SAA). Increases in blood levels of these factors correlate with, and are diagnostic of, an increased state of inflammation [13,14,15]. To measure the levels of these factors, we made use of plasma samples that were obtained 33 weeks after initiation of the experimental diets. We found that the levels of haptoglobin and SAA were significantly lower in the circulation of mice undergoing IMR as compared with the corresponding levels in control-fed mice (Fig. 1C-D). To further explore the state of inflammation in high-fat diet-fed mice, we also measured the circulating levels of interleukin-6 (IL-6). Serum levels of IL-6 have been shown to correlate with inflammation in patients with psoriatic dermatitis [16]. We observed a trend towards lower levels of IL-6 in mice undergoing IMR as compared with controls (Fig. 1E), although this difference was not significant.
We also assessed the state of inflammation in animals subjected to both diets by measuring leukocyte numbers in whole blood samples from 6-month-old male mice that had been maintained on either the CF or IMR regimens for 16 weeks (Fig. 1F and Supplemental Table 1). An elevated white blood cell (WBC) count of ~ 10,000/µl or greater (otherwise known as leukocytosis) is typically indicative of an increased state of inflammation in infection-free animals [17, 18]. As expected, we observed that control-fed mice demonstrated an average leukocyte count consistent with this condition (9,844/µl). Interestingly, IMR resulted in a lower average cell count (8,228/µl), although this difference was not significant. It is possible that a longer diet duration, such as the 33 weeks used for measurement of the acute phase reactants, might result in a significant and/or larger difference between the WBC counts observed for control-fed and intermittently methionine-restricted mice. However, such samples were utilized in their entirety for other analyses. In any case, when taken together, the above results are consistent with IMR conferring a moderate protection against systemic inflammation and the development of ulcerative dermatitis.
Selenium supplementation protects high-fat diet-fed mice against dermatitis and chronic inflammationSimilar experiments to those described above were also performed for mice consuming a selenium-supplemented diet. To assess the effects of selenium supplementation on dermatitis and inflammation, male mice were continuously maintained on one of two high-fat diets, these being either (1) a methionine-replete control diet or (2) an otherwise identical diet containing 0.0073% sodium selenite (Supplemental Table 1). Both diets were initiated when animals were 8 weeks of age and continued until they displayed evidence of dermatitis, at which point high-fat diet feeding was discontinued. For those animals that failed to develop dermatitis, high-fat diet feeding was discontinued after 90 weeks. As expected, nearly all control-fed animals developed dermatitis (Fig. 2A and Fig. S1), with a median dermatitis-free survival duration nearly identical to that observed for the control mice in the IMR experiment (327 days vs. 341 days; Figs. 1B and 2B). However, mice consuming a selenium-supplemented diet demonstrated a dramatic delay in the development of dermatitis, with a 292 day increase in median dermatitis-free survival (619 days vs. 327 days; p = 0.0112). Interestingly, by the conclusion of the experiment (i.e., after 90 weeks of feeding), all control-fed animals but one had developed dermatitis, whereas only 50% of selenium-supplemented mice had ever demonstrated evidence of this condition.
Fig. 2
Selenium supplementation protects high-fat diet-fed mice against dermatitis and chronic inflammation. Shown are (A) representative images of mice that were either control-fed (CF) or selenium-supplemented (CF + SS), as well as (B) the longitudinal incidence of dermatitis for such animals. The average plasma levels of the inflammation markers (C) haptoglobin and (D) serum amyloid A (SAA) in animals fed the indicated diets are also provided, as well as those of the pro-inflammatory cytokine (E) interleukin-6 (IL-6). As a further assessment of systemic inflammation in these animals, white blood cell (WBC) concentrations were also determined (F). N = 8 for all groups. For panel B, the difference between treatment groups is statistically significant (p = 0.0112). For panels C-F, bars denote SEM and statistically significant differences are indicated (*, p < 0.05; **, p < 0.01)
To test whether, similar to the results for IMR, the protection against dermatitis conferred by selenium supplementation correlated with reduced systemic inflammation, we measured the circulating levels of the acute phase reactants. For animals maintained on the experimental diets for 34 weeks, we found the plasma levels of haptoglobin and SAA to be significantly lower in mice undergoing selenium supplementation as compared with the corresponding levels in control-fed mice (Fig. 2C-D). We also measured the circulating levels of IL-6 in mice fed both diets and observed a trend towards lower levels in mice undergoing selenium supplementation (Fig. 2E), although this difference was not significant.
As a final test of inflammation in animals fed the two diets, we tested the extent of leukocytosis (if any) in both selenium-supplemented and control-fed mice. For this analysis, whole blood samples were obtained from mice 47 weeks after initiation of the experimental diets. Similar to the results from the IMR experiment (Fig. 1F), control-fed mice were found to have a WBC count indicative of leukocytosis (11,700/µl; Fig. 2F). However, selenium supplementation resulted in a significantly lower average cell count of 5,743/µl, a value that is well within the normal range (2,000–10,000/µl). Taken together, the above results demonstrate that selenium supplementation confers a robust protection against both systemic inflammation and the development of dermatitis.
Comments (0)