
Remember me




A 20-year-old female was referred to hematology center for CAR-T cell therapy following relapse of Philadelphia-negative B-cell acute lymphoblastic leukemia (ALL). The patient was initially diagnosed with ALL in March 2022 at another institution after presenting with lower limb bruising and marked leukocytosis (WBC 63,710/mm³) with 80% peripheral blood blasts. She received standard multi-agent chemotherapy (LAL1913 protocol), which included an initial steroid and cyclophosphamide prephase, followed by induction therapy with vincristine, idarubicin, peg-asparaginase, and dexamethasone, along with prophylactic intrathecal methotrexate, cytarabine, and prednisone until April 2022. The patient subsequently underwent seven additional cycles of induction and consolidation therapy until December 2022, achieving measurable residual disease (MRD) negativity. Maintenance therapy began in January 2023 and continued until January 2024, when she experienced her first relapse.
Upon relapse confirmation (85% blasts on bone marrow morphology; CD22 positivity confirmed by flow cytometry), the patient received a single cycle of Inotuzumab as a bridge to CAR-T, once again achieving MRD negativity by flow cytometry. Following leukapheresis for CAR-T manufacturing, a second Inotuzumab cycle was administered, with MRD negativity confirmed via flow cytometry and molecular testing. The patient received CAR-T cell infusion in April 2024, following standard lymphodepleting chemotherapy with fludarabine and cyclophosphamide. Her post-CAR-T course was uneventful, and she remained in molecular remission until July 2024, when MRD re-emerged (10⁻³).
In response to molecular relapse, she was started on blinatumomab therapy, which was initially complicated by neurotoxicity. Treatment was temporarily halted, and the patient received antiepileptic therapy and high-dose corticosteroids. Blinatumomab was then resumed on an outpatient basis as a bridge to transplant, along with intrathecal chemotherapy, steroids, and weekly vincristine. This outpatient period was further complicated by SARS-CoV-2 infection, treated with nirmatrelvir.
The patient was admitted for allogeneic hematopoietic stem cell transplantation in early October 2024. A bone marrow aspirate at admission confirmed morphologic persistence of disease, prompting the administration of vincristine and corticosteroids. Due to persistent SARS-CoV-2 positivity on day 9 of admission, she received a course of remdesivir based on infectious disease consultation. Admission microbiological testing for gastrointestinal pathogens (bacteria, viruses, and parasites), fungi (germ tube test), multidrug-resistant organisms, and cytomegalovirus was negative.
Conditioning therapy commenced on day 26 and included total body irradiation (2 Gy twice daily on days − 8 to -6), fludarabine (40 mg/m² on days − 5 to -3), followed by infusion of peripheral blood stem cells from a 7/8 HLA-mismatched unrelated donor. Graft-versus-host disease (GvHD) prophylaxis included anti-thymocyte globulin (ATG), post-transplant cyclophosphamide (50 mg/kg on days + 3 and + 5), cyclosporine (from day + 5), and mycophenolate mofetil (1000 mg twice daily from day + 5 to + 30). A summary of the clinical course is presented in Fig. 1a.
Fig. 1
Schematic timeline reporting information on the clinical history of the patient (a). Patient’s blood inflammatory markers and microbiological findings during the period November 2024-January 2025 (b). Panel shows haemato-chemical inflammatory markers sepsis-related: white blood cells (WBC), C-reactive protein (CRP), procalcitonin (PCT). Laboratory microbiological findings regard bacterial, fungal and viral pathogens. Abbreviation: WBC: white blood cells; C-reactive protein (CRP); procalcitonin (PCT); RS: Rectal Swab; BAL: Broncho-Alveolar Lavage
Post-transplant complications included pulmonary aspergillosis (treated with isavuconazole and caspofungin) and HHV-6 encephalopathy (treated with foscarnet). Rhinovirus was also detected. Febrile neutropenia and catheter-related bloodstream infection (BSI) due to Staphylococcus epidermidis occurred during conditioning and were treated with meropenem, vancomycin, and liposomal amphotericin B. An Enterococcus faecalis infection was treated with cefepime, which was later discontinued due to neurotoxicity. Microbiological findings and inflammatory markers are presented in Fig. 1b.
In January 2025, the patient experienced additional severe transplant-related complications, including acute and chronic GvHD (maximum grade 3 cutaneous and gastrointestinal), for which she received corticosteroids, ruxolitinib, octreotide, and biweekly therapeutic plasma exchange. She also developed sinusoidal obstruction syndrome (treated with defibrotide) and transplant-associated thrombotic microangiopathy (TA-TMA), for which she received fresh frozen plasma infusions. Despite maximal therapeutic and supportive interventions, the patient died of multiorgan toxicity on day + 76 post-transplant. Beginning on day + 70 of admission, blood cultures drawn from peripheral veins, peripherally inserted central catheter (PICC), central venous catheter (CVC), and midline catheter were all positive for Lacticaseibacillus rhamnosus (L. rhamnosus). During this time, inflammatory markers associated with sepsis, including D-dimer and procalcitonin, were markedly elevated (Fig. 1b). On day + 70, aerobic and anaerobic blood culture bottles were incubated using the BACT/ALERT® 3D system (bioMérieux, Marcy l’Étoile, France). Three aerobic cultures (one from PICC, two from CVC) became positive after 46 h, revealing Gram-positive rod-shaped bacteria. Identification using the MALDI BioTyper® system (Bruker Daltonics, Bremen, Germany) confirmed L. rhamnosus with a score of 2.38 (reliable identification: >1.699). The second and third best matches also identified L. rhamnosus (scores: 2.31). Positive cultures were subcultured on Tryptic Soy Agar (TSA) and Columbia agar and incubated at 37 °C with 5% CO₂, confirming the identification.
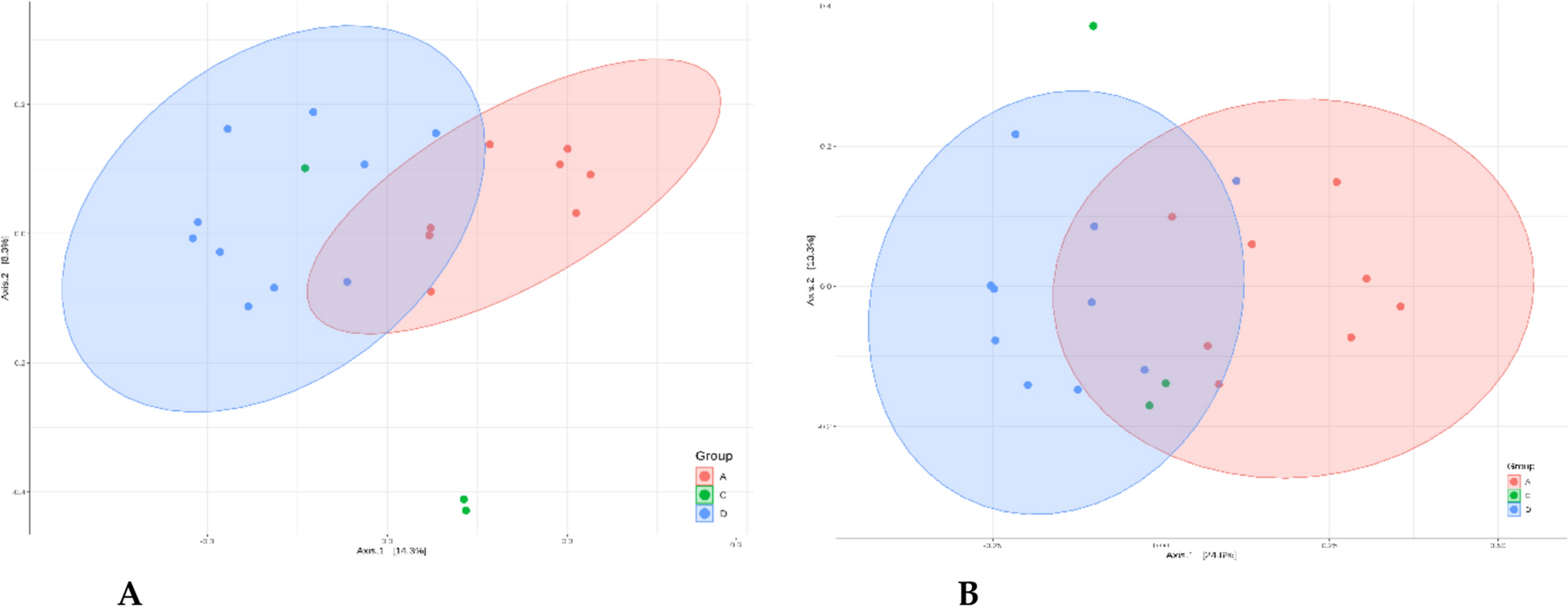
The genome of the clinical isolate was sequenced (see Additional File 1: Methods). Upon detection of L. rhamnosus in blood cultures, a faecal sample was collected and subjected to 16 S rRNA gene-targeted gut microbiota profiling (see Additional File 1: Methods). As shown in Fig. 2a, sequencing of the V3-V4-V6 regions revealed highest relative abundances for L. paracasei (42.4%), Pediococcus acidilactici (18.7%), and unclassified Lactobacillaceae (16.8%). Given that 16 S sequencing cannot reliably differentiate species within the Lactobacillus casei group, shotgun metagenomic sequencing was performed. This analysis revealed L. rhamnosus as the dominant species (71.6%), followed by Pediococcus acidilactici (17.4%) (Fig. 2b). Taxonomically assigned reads corresponding to L. rhamnosus were extracted and assembled for genomic comparison with the clinical blood isolate. The two reconstructed genomes differed by only 18 single nucleotide polymorphisms (SNPs), strongly suggesting a clonal relationship. These findings demonstrate gut domination by L. rhamnosus and subsequent bloodstream infection, indicating a potential translocation event and highlighting the importance of intestinal microbiota surveillance in high-risk patients.
Fig. 2
Gut Microbiota characterization at the moment of L. rhamnosus isolation from blood cultures. Panel a shows 16 S rRNA gene-targeted intestinal microbiota profiling by using V3-V4-V6 hypervariable regions, while panel b shows shotgun metagenomics results
Discussion and conclusionsClinical management of immunocompromised patients, in particular hemopoietic cell transplant patients, remains critical, due to their increased susceptibility related to long-term hospitalization and consequent enhanced exposure to multi-drug-resistant bacteria [2, 32] and opportunistic pathogens [1, 12]. In our patient, severe impairment of immune functions may have contributed to L. rhamnosus BSI with an unfavourable outcome.
Risk factors in our patient included the acute hematologic malignancy, prior treatment, included B cell-depleting therapy, which directly diminishes host immune defences [1], and extensive use of antibiotics, which promotes bacterial colonization and infection. Together, these factors resulted in dysbiosis of the gut microbiota, favouring the overgrowth of pathobionts [32, 33]. Ultimately, intestinal domination occurred, resulting in occupation of the microbiota by a single predominating bacterial taxon [3], decreasing microbial diversity and increasing the risk of bacterial translocation into the bloodstream [12].
In this study, we demonstrated, for the first time, experimental evidence of intestinal domination by L. rhamnsosus, which was then rapidly and directly identified from blood specimens [34]. This case demonstrates the role of gut microbiota analysis in high-risk patients, as initial bacterial colonization can give rise to subsequent BSI.
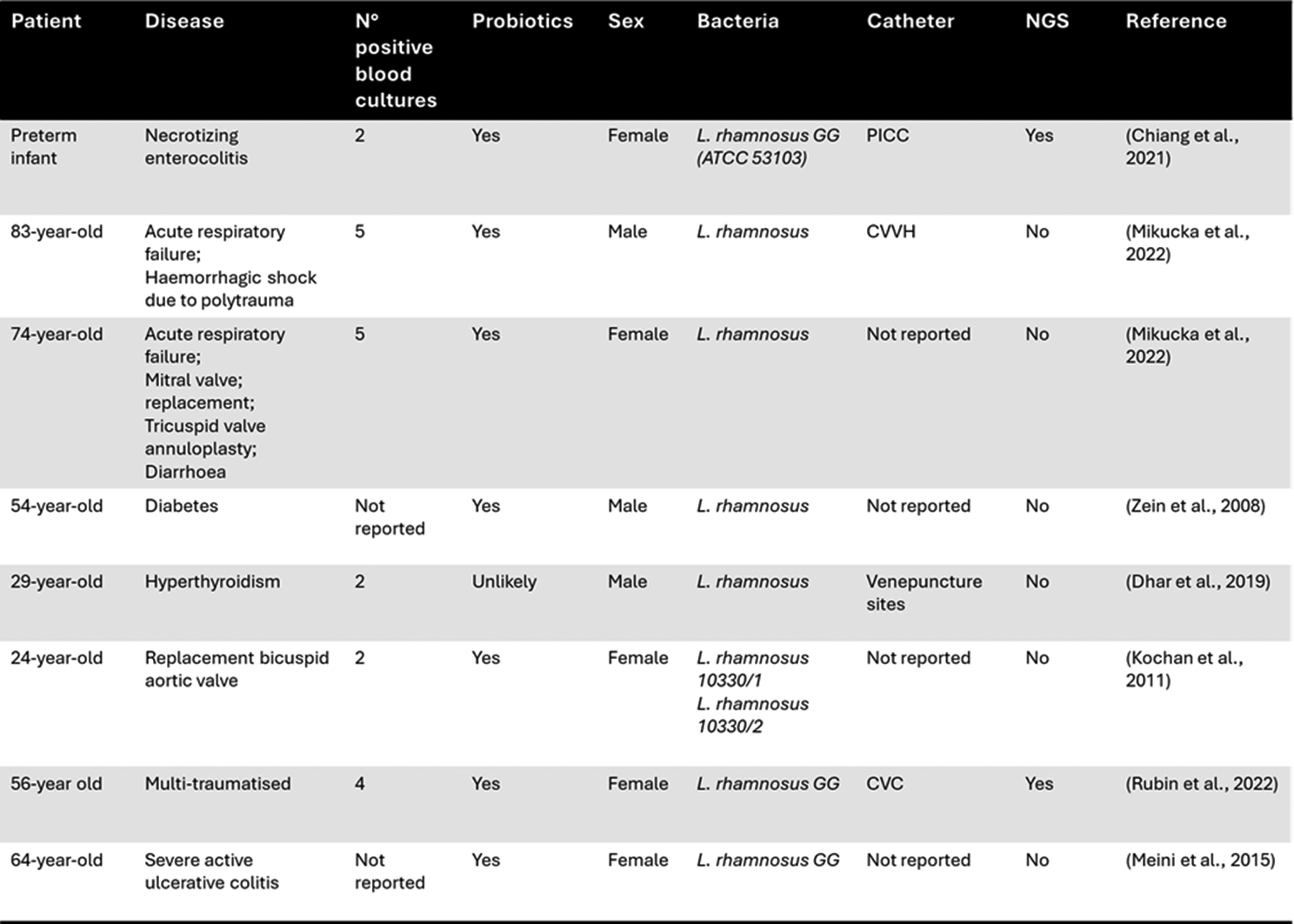
A number of clinical cases on pathobiont-associated BSIs, including L. rhamnosus, have been reported, as shown in Table 1 [17, 35]. Although Lactobacillus-dominant intestinal microbiota has been reported in some individuals, it remains relatively uncommon in the general population and it may be associated with a specific dietary pattern, probiotic use, or underlying clinical conditions, including polytrauma, severe ulcerative colitis, or hyperthyroidism. This study is the first to show pathobiont-associated BSI mediated by Lactobacilli mediated intestinal domination a condition that is not common. Unfortunately, data on intestinal microbiota composition in Italy are limited, even though in European countries, and not only, Enterococcus or multi-drug-resistant Gram-negative bacteria intestinal domination is more common [3, 36].
Of note, the use of probiotic supplements and the intake of fermented foods containing Lactobacillus (e.g., yogurt, cheese, kefir) were not reported prior to the onset of BSI suggesting that genetic, environmental or metabolic factors may have affected gut microbiome. Interestingly, many of these factors represent important confounding factors to delineate both healthy and non-healthy gut microbiome status [16].
Our case shows that monitoring the state of eubiosis may be relevant in hematologic patients, who are subject to extensive use of the antibiotics as well as administration of probiotics, including L. rhamnosus [21, 37]. Indeed, probiotics can interact with host immune cells in the mucosal epithelium, modulating cytokine secretion and causing systemic effects [38]. Lactobacilli can activate macrophages to secrete inflammatory cytokines such as Interleukin-6 (IL-6) and Tumor Necrosis Factor-α [39]. Probiotics can positively modulate B cell activity, as stimulating IL-6 release causes B cell clonal expansion. Some probiotics species may enhance Interleukin-12 production, stimulating development of T-cells and increased NK cell reactivity [38, 40]. Thus, probiotic administration may potentially connect innate and acquired immunity by multiple pathways, though the precise molecular mechanisms remain to be determined. We hypothesize that an overgrowth of these species may induce a significant inflammatory state, which contributes to significant modifications of the host immune response together along with anti-neoplastic treatments [1, 3, 41], and use of other agents, including prophylactic treatment with TMP/SMX [42]. On the other hand, several evidence suggest that COVID-19 may induce significant alterations in the gut microbiota, including a decrease in microbial diversity and an increase in opportunistic pathogens, which can persist even after the acute phase of infection has resolved [43,44,45]. The loss of the normal equilibrium formed by Firmicutes and Bacteroidota [46] may predispose to bacterial translocation across the intestinal barrier [47].
Therefore, we acknowledge the potential role of prior COVID-19 infection as a contributing factor to the patient’s dysbiosis, in addition to the impact of transplantation-related variables. Lactobacillus BSIs are intrinsically resistance to glycopeptides, including teicoplanin and vancomycin [35], even though European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints are not available for Lactobacilli [48].
Intriguingly, other probiotic genera, such as Bifidobacterium, generally exhibit lower levels of intrinsic resistance to antibiotics and have fewer reported associations with BSIs. For instance, Bifidobacterium species show intrinsic resistance to glycopeptides like vancomycin, like Lactobacillus, but typically lack acquired resistance genes such as vanA, vanB, or vanC [49,50,51]. Additionally, novel probiotics like Akkermansia muciniphila also displays intrinsic resistance to glycopeptides [51], suggesting that antimicrobial resistance profiles may have significant clinical implications in the management of gut microbiota.
However, potential risk factors specifically for Lactobacillus BSIs are difficult to identify: a differential time to positivity between blood cultures obtained from CVC and peripheral vein of about four hours suggests the gut as the source of the BSI, in spite of the potential for Lactobacillus ability to form biofilm on catheter lumina [35]. In our study, we were able to demonstrate L. rhamnsosus intestinal domination, thanks to a complementary analysis of the gut microbiota (metagenomics) together with whole genome sequencing of the clinical isolate.
Lactobacilli may behave as an opportunistic pathogen [23]: several studies have warned against the possible danger of L. rhamnsosus, which has often been given as a probiotic treatment for intestinal disorders [21]. Their inherent antibiotic resistance profile may selectively promote their growth with respect to other microbial components.
While metagenomic approaches are well established in comparative studies involving two or more groups, their integration into diagnostic workflows remains controversial, largely due to the lack of standardized procedures, normalization strategies, and validated pipelines, particularly in the context of single-sample analyses and biomarkers discovery [16].
We acknowledge the limitations of this study, including the absence of baseline samples which preclude inference of causality regarding the origin of intestinal domination. Nonetheless, our findings join to the growing evidence highlighting a temporal association between intestinal domination and BSI. This case supports the concept that gut microbiota analysis, including relative abundance profiling, could help to identify taxa that may pose a risk for bloodstream infection in high-risk patients, especially those undergoing prolonged hospitalization, broad-spectrum antibiotic therapy, and potential exposure to prebiotic or probiotic interventions, all of which may predispose to the overgrowth of pathobionts.
While sequencing-based approaches (both 16s metabarcoding and shotgun metagenomics) may not yet be practical for a routine clinical use, they offer a comprehensive tool for detecting relevant microbial shifts and for informing the development of faster and accurate targeted assays for early risk stratification and infection surveillance. Notably, many pathobionts are not associated with specific disease syndromes [52] or may only exert a transient impact on the microbiome [16]. As such, they often do not elicit a targeted clinical suspicion and are not routinely included among the etiological agents covered by standard diagnostic panels. In this context, microbiome findings could play a pivotal role in the development of novel diagnostic assays based on quantitative PCR (both real-time and digital PCR). Translation of the metagenomic findings may be essential for the successful integration of these technologies in diagnostic workflows no less than their ability to outperform current diagnostic methods.
Comments (0)