
Remember me

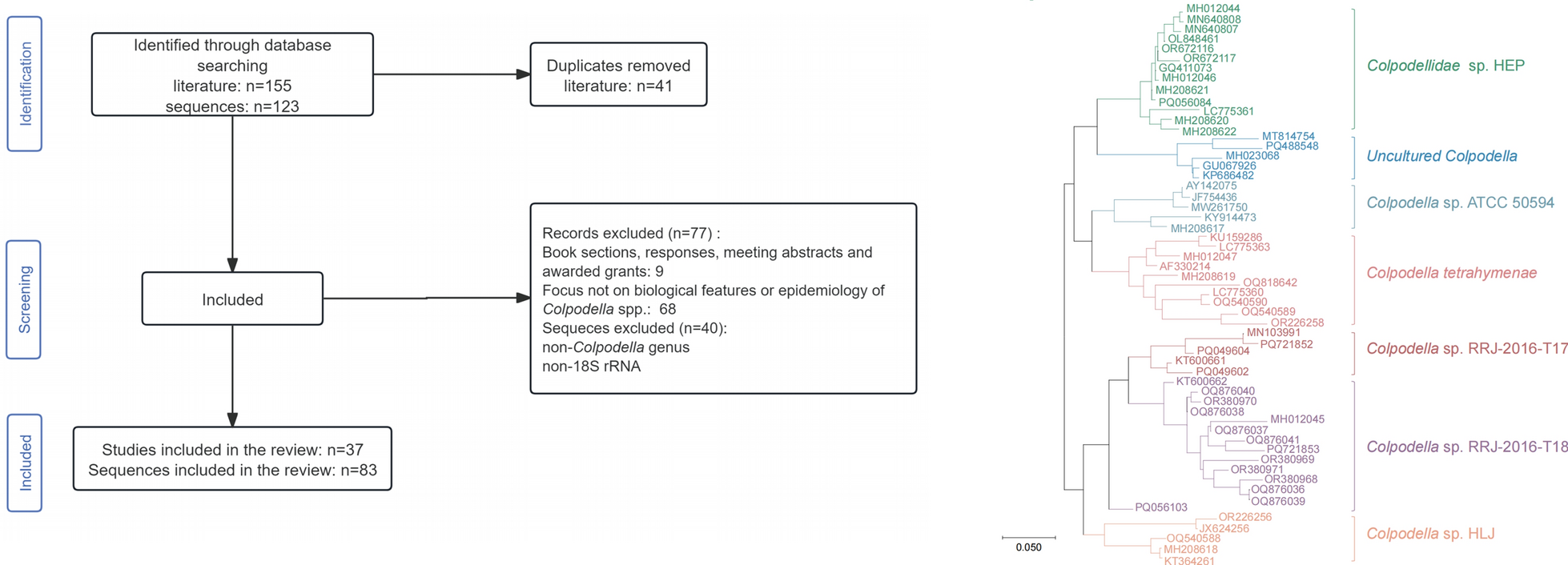

The systematic search identified 155 records, with 41 duplicate records removed. Following full-text review, 77 records were excluded according to the inclusion criteria. Ultimately, 37 articles met the inclusion criteria (Fig. 1). For genetic analysis, a total of 123 sequences were retrieved, and 83 sequences were included in the analysis based on the exclusion criteria (Fig. 2).
Fig. 1
Flowchart of the literature screening process
Fig. 2
Flowchart of the sequences screening process
Biological characteristics of Colpodella spp.Taxonomic status and species of Colpodella spp.Colpodella spp. are a small group of free-living predatory protozoans belonging to the the domain Eukaryota, order Colpodellida, family Colpodellaceae, and genus Colpodella. They primarily feed on protozoa and algae, have occasionally been identified in vertebrate hosts and arthropod vectors, and are phylogenetically closely related to apicomplexan protozoa, which comprise a diverse group of specialized intracellular parasites [6, 13, 14], including Plasmodium, Toxoplasma, and Cryptosporidium. Currently, Colpodella spp. include the formally described species Colpodella tetrahymenae and Colpodella angusta, as well as at least 11 unclassified and unnamed species in GenBank (Additional file 3: Table S3).
Morphological features of Colpodella spp.Similar to other apicomplexan protozoa, Colpodella spp. possess micropores and apical complexes, which comprises rhoptries, micronemes, pseudo-conoids, polar rings and subpellicular microtubules [6, 15]. In contrast to the complete ring conoid of the apicomplexan protozoa, the pseudo-conoid of Colpodella spp. consists of an incomplete ring of microtubules [16]. The number of microtubules comprising the pseudo-conoids varies among Colpodella species. C. angust generally has 20–27 microtubules while C. tetrahymenae has only 4–5 [17].
Life cycle of Colpodella spp.The in vitro life cycle of Colpodella spp. comprises trophozoite stage and cyst stage. Traditional techniques (wet mounts and Giemsa-stained smears) enable observation of trophozoite movement and cyst detection [18, 19]. However, these methods can not distinguish between Colpodella sp. and prey cysts. Sam-Yellowe TY et al. [20] developed a novel trichrome staining technique that enabled detailed differentiation and observation of Colpodella sp. cyst stages.
The trophozoite stage is the active feeding phase of Colpodella spp. After constructing tubular connections to attach to prey, trophozoites suck its cytoplasm. After feeding, the anterior end of the trophozoite disintegrates, accompanied by the disappearance of flagella and organelles. The food vacuole, along with the remaining cytoplasm and nucleus, forms a pre-cyst, which subsequently differentiates into an early demilune cyst and initiates encystation [16, 21]. Most cysts of Colpodella spp. typically contain two or four juvenile trophozoites [6, 15, 16]. However, Colpodella vorax and Colpodella sp. ATCC 50594 exhibit cysts containing up to 10 juvenile trophozoites [16, 22]. Trophozoites emerge from cysts, appearing either as individual cells or as paired, resuming feeding activities to complete the life cycle. Colpodella sp. ATCC 50594 has a 36-h life cycle, with peak activity occurring between 20 and 28 h [21].
Mechanisms of nutrient uptake of Colpodella spp.Colpodella spp. utilize myzocytosis for feeding. Myzocytosis, a primarily predatory trophic mode, which is based on penetration of the prey surface and sucking of its contents via specialised organelles morphologically similar to those used by the apicomplexan zoites for invasion [23]. Following attachment of the pseudo-conoids of Colpodella spp. trophozoites to the prey surface, the microtubules at the attachment site dilate, enabling the engulfment of the prey's cell membrane and cytoskeleton. This process establishes a specialized tubular connection between predator and prey, enabling directional transport of prey cytoplasm into the posterior food vacuole. At the end of feeding, Colpodella spp. detach from the prey surface and continue to search for new prey [17, 21, 22]. The most common predator–prey interaction involves the attachment of a single Colpodella individual to a single prey organism during feeding. In addition, instances of two to three Colpodella individuals simultaneously attaching to a single prey cell have been observed [17, 21]. Unlike pathogenic apicomplexan parasites, which form parasitophorous vacuol by invading host cells during feeding, Colpodella spp. form food vacuoles without invading their prey. Furthermore, micropores, which are present in all apicomplexan parasites, are hypothesized to function in endocytosis. In Toxoplasma gondii, the micropore has been identified as an essential organelle for nutrient uptake via endocytosis [24]. Except for Colpodella sp. ATCC 50594, endocytosis has not been reported in other Colpodella species [25]. Therefore, it remains unclear whether Colpodella spp. employ endocytosis as an additional feeding mechanism alongside myzocytosis. Colpodella species may utilise endocytosis to absorb and survive on nutrients in human, animal, and arthropod tissues. Alternatively, they may engage in contact-dependent interactions with cells, leading to cell and tissue destruction or invasion of human cells [26, 27].
The key role of apical complexes-related proteinActin plays a critical role in the invasion of host cells by apicomplexan parasites, such as T. gondii and Plasmodium falciparum [28, 29]. Cytochalasin D blocks the action of actin in T. gondii [30]. Similarly, Cytochalasin D treatment of Colpodella sp. ATCC 50594 induces cytoskeletal deformation and disrupts the tubular tether during predation, indicating that actin is involved in tubular tether formation [31]. In addition, Apical complexes organelles use their related proteins to initiate host cell invasion and facilitate intracellular trophozoite survival [32]. Colpodella spp. also have an apical complex. Sam-Yellowe TY et al. demonstrated that antibodies to proteins involved in host cell invasion by apicomplexan parasites cross-reacted with apical proteins of Colpodella sp. ATCC 50594. Antibodies targeting the rhoptry protein RhopH3 of the Plasmodium species cross-react with the tubular connection structures formed between Colpodella sp. ATCC 50594 and its prey during the myzocytosis stage [33]. Similarly, antibodies against apicomplexan invasion-related proteins, such as inner membrane complex subunit 3 (IMC3), apical membrane antigen 1 (AMA-1), erythrocyte-binding antigen 175 (EBA-175), and Plasmepsin II, also showed cross-reactivity with this organism [21]. The cross reactivity of antibodies against apical complex proteins with Colpodella sp. ATCC 50594 proteins suggests that events of myzocytosis may have preceded events that led to zoite internalization within host cells in intracellular parasitism [21]. These findings suggest that the function of Colpodella spp. apical proteins may resemble that of apicomplexan parasites during host cell invasion, indicating that Colpodella spp. may interact with host cells through their proteins, potentially causing cellular damage and facilitating invasion.
Both feeding mechanism and the structure and function of their apical complex organelles of Colpodella spp. show significant similarities to those involved in host cell invasion by apicomplexan parasites. However, there is still a lack of research on the invasion mechanisms of Colpodella spp., even though Yuan et al. have observed Colpodellidae sp. HEP in human red blood cells [7].
Epidemiological studies on Colpodella spp.Human infections with Colpodella spp.To date, Colpodella spp. have been identified in three patients (Table 1). The first case was reported in 2008 in Yunnan Province, China, who was a 57-year-old woman, presented with cough and malaise for 6 months and was found to have several hematological abnormalities. Microscopic examination for peripheral blood smear identified parasites resembling Babesia spp. Then she was treated with atovaquone and azithromycin. Retrospective PCR targeting the 18S rRNA gene and sequence analysis revealed she was infected with Colpodella sp. which had an 89.0% similarity with C. tetrahymenae, and was designated as Colpodellidae sp. HEP [7]. In 2018, our research team reported the second case of Colpodella infection, a 55-year-old female patient with a confirmed history of tick bite in Heilongjiang Province, China. Neurological examination revealed moderate nuchal rigidity. The initial clinical diagnosis was Lyme disease. Administration of doxycycline led to improvement in the patient’s condition. However, retrospective testing of blood and cerebrospinal fluid samples was negative for all known tick-borne pathogens. Unexpectedly, the recovered 18S rRNA sequence from her cerebrospinal fluid sample was closely related to Colpodella spp. (89.0–90.0% similarity) and was provisionally designated as Colpodella sp. HLJ [8]. Notably, in 2021, Colpodella sp. was identified in urine samples from a 70-year-old female patient with lung disease in Romania using wet mounts and Giemsa-stained smears. The patient presented with breathing difficulties and multiple chronic diseases, but exhibited no urinary pathology or symptoms before or during hospitalization. Treatment included ceftriaxone and metronidazole. Final urine sediment examinations prior to discharge confirmed the absence of the parasite [9].
Table 1 Description of human cases of Colpodella spp. infectionThese three cases suggest that Colpodella spp. may act as opportunistic pathogens in humans. However, as these cases were identified retrospectively, the target organs, pathogenicity, sources of infection, and transmission mechanisms remain poorly characterized. Although Colpodella sp. HLJ was initially associated with a tick bite in the patient, retrospective analysis of tick-derived sequences revealed significant divergence from the human isolate.
Genetic diversity of Colpodella spp.Despite the limited number of reported human infections, the genetic diversity of Colpodella spp. among animals and vectors has been increasingly recognized through phylogenetic analyses. Understanding the genetic variation within this genus is crucial for elucidating its potential pathogenicity, host adaptation, and transmission dynamics.
A phylogenetic tree was constructed using all available 18S rRNA sequences of Colpodella spp. retrieved from GenBank (Fig. 3). The phylogenetic tree was divided into seven distinct clades based on sequence similarity (> 87.0%) between the most closely related sequences in adjacent branches. Species classification was conducted using a similarity threshold of 95% provisionally, with each of the seven clades containing multiple Colpodella species (Additional File 2: Table S2). Representative Colpodella species were provisionally named for each clade, including Colpodellidae sp. HEP., Uncultured Colpodella, Colpodella sp. ATCC 50594, Colpodella tetrahymenae, Colpodella sp. RRJ-2016-T17, Colpodella sp. RRJ-2016-T18, Colpodella sp. HLJ. To date, only two Colpodella strains, Colpodellidae sp. HEP (accession numbers: GQ411073) and Colpodella sp. HLJ (accession numbers: KT364261), have been associated with human infections, while nearly 15 strains are linked to animals and vectors. The hosts of Colpodella spp. in clades of Colpodellidae sp. HEP and Colpodella sp. HLJ include wildlife, livestock, ticks, pets and humans, suggesting a high potential for an animal-vector-human transmission cycle. Although the clades of Colpodella sp. ATCC 50594, Colpodella sp. RRJ-2016-T17, Colpodella sp. RRJ-2016-T18 and Colpodella tetrahymenae have not been found to be directly associated with human infections, the host ranges of these species encompass wildlife, pets, livestock, and vector insects such as ticks and flies. This wide host distribution suggests a potential risk of transmission to humans. In addition, the clade of Uncultured Colpodella is currently only associated with livestock, with additional environmental detections..
Fig. 3
Phylogeny of Colpodella spp. sequences of 18S rRNA gene, using the Neighbor-Joining method with Bootstrap 1000 replicates. The phylogenetic tree was constructed using the MEGA 11.0 software. Sequences from the same institution, at the same time, and from the same source are considered to be from the same species if their similarity is > 99% [47], and only one is shown in the figure. For specific selection reasons, please refer to Table S3
Geographical distribution of Colpodella spp.Over the past two decades, Colpodella spp. have been identified to be widely distributed across multiple continents-Asia, Europe, the Americas, and Africa, inhabiting diverse geographical regions and ecological environments (Additional file 4: Table S4). Colpodella species currently exhibit a broad geographic distribution, with all six clades (excluding the Uncultured Colpodella clade) reported across Asia. The Colpodella sp. HLJ clade currently exhibits a relatively limited geographical distribution pattern, mainly distributed in Heilongjiang Province, Yunnan Province, Shandong Province and Guizhou Province in China. Notably, the Colpodellidae sp. HEP clade has been detected in both Asia and Africa, while clades of Colpodella sp. RRJ-2016-T17 and Colpodella sp. RRJ-2016-T18 show transcontinental distributions, occurring in Europe—with the latter additionally present in Africa. Most remarkably, the clades containing Colpodella sp. ATCC 50594 and C. tetrahymenae demonstrate the widest distribution range, spanning Africa, Asia, and the Americas. The widespread distribution of Colpodella spp. suggests not only a broad ecological niche but also a strong environmental adaptability [14, 34, 35]. These characteristics together promote their potential for cross-regional spread.
Risk of human infection with Colpodella spp.First, the widespread distribution of Colpodella spp. further increase their likelihood of infecting humans. Colpodella spp. were frequently detected not only in hosts and vectors, but also in a wide range of environmental samples. For example, Colpodella sp. ATCC 50594 was originally isolated from brown woodland soil samples in Gambrill State Park, Maryland, United States [14]. In addition, Colpodella spp. have also been identified in specific reef environments, including coral tissue, mucus, and associated seawater sediments [35]. Similarly, Colpodella spp. were also detected in water samples collected from small ditches near a tiger park in China [34]. These findings suggest that Colpodella spp. may also be transmitted through environmental media such as soil and water bodies.
Second, the extensive prevalence of Colpodella spp. within zoo animals (e.g., tigers), domestic animals (e.g., horses), pets (e.g., dogs and cats) and wildlife (e.g., foxes and birds) in intimate contact with humans substantially escalates the likelihood of human infection. In China, Colpodella spp. have been detected in fecal samples from different felines (Panthera tigris, Panthera leo, Panthera onca, Lynx lynx) from Harbin Zoo, Heilongjiang Province [36], blood from horses in Ordos, Inner Mongolia [37], pet cats and dogs in Guizhou Province [38]. A South China tiger (Panthera tigris amoyensis) in Fujian Province was bitten by ticks and exhibited clinical symptoms such as severe jaundice throughout the body, enlarged liver and spleen, and bleeding in the kidneys and lymph nodes [34]. The sequence of Colpodella sp. detected in the blood of the South China tiger showed 90.1% similarity to Colpodellidae sp. HEP and 90.4% similarity to Colpodella sp. HLJ. Given the close contact between humans and these animals, the risk of infection with Colpodella spp. is significantly increased.
Humans can become infected with Colpodella spp. not only through daily contact with domestic animals and pets but also through increasing interactions with wildlife. In Cyprus, Colpodella spp. were identified in fecal samples from Cyprus red foxes (Vulpes vulpes indutus), Eurasian coots (Fulica atra) and ducks (Anas sp.) [39].
Third, blood-sucking ticks and flies, as important vectors, will play a significant role in the transmission of Colpodella spp. among people. The patient, from Heilongjiang Province, China, had a history of tick bite. During field investigations in the woodlands surrounding the patient's living area in Heilongjiang Province, 474 host-seeking adult Ixodes persulcatus ticks were screened, among which two were identified as positive for Colpodella spp. [8]. In China, different Colpodella spp. were also detected in other tick species: ticks (Dermacentor nuttalli, Dermacentor abaensis, Haemaphysalis qinghaiensis) across nine counties in Qinghai Province [40], goat-attached Haemaphysalis longicornis ticks in Shandong Province [41], Amblyomma javanense ticks on Malayan pangolins intercepted by customs in Guangdong Province [42] and ticks (Haemaphysalis spp., Dermacentor spp., Rhipicephalus spp.) around a tiger park, in Fujian Province [34], further suggesting that ticks may act as their vectors. In addition to findings in China, Colpodella spp. were also identified in Hyalomma dromedarii ticks infesting camels in southern Egypt [43], as well as in Rhipicephalus microplus ticks that feed on cattle in Mozambique [44]. Moreover, this microorganism has been detected in Stomoxys indicus flies that are associated with horses in Thailand [45]. Strong associations between Colpodella spp. and multiple tick species and other arthropods suggest that they may serve as key vectors for their transmission and spread.
Through a thorough review of literature on the distribution of Colpodella spp.in the environment, animals and vectors (Table 2), we demonstrated the wide geographical distribution and host range of Colpodella spp. Potential transmission routes for human infection with Colpodella spp. may include vector-borne transmission and indirect contact transmission. Being bitten by infected vectors or coming into contact with contaminated objects can lead to infection Colpodella spp. In addition, Colpodella spp. have also been detected in animal feces in current studies [36, 39], indicating that fecal–oral transmission may also exist. These findings highlight the complexity of their transmission routes and pose a potential threat to public health that cannot be ignored.
Table 2 Carriage records of Colpodella spp. in animals and arthropod vectors
Comments (0)