
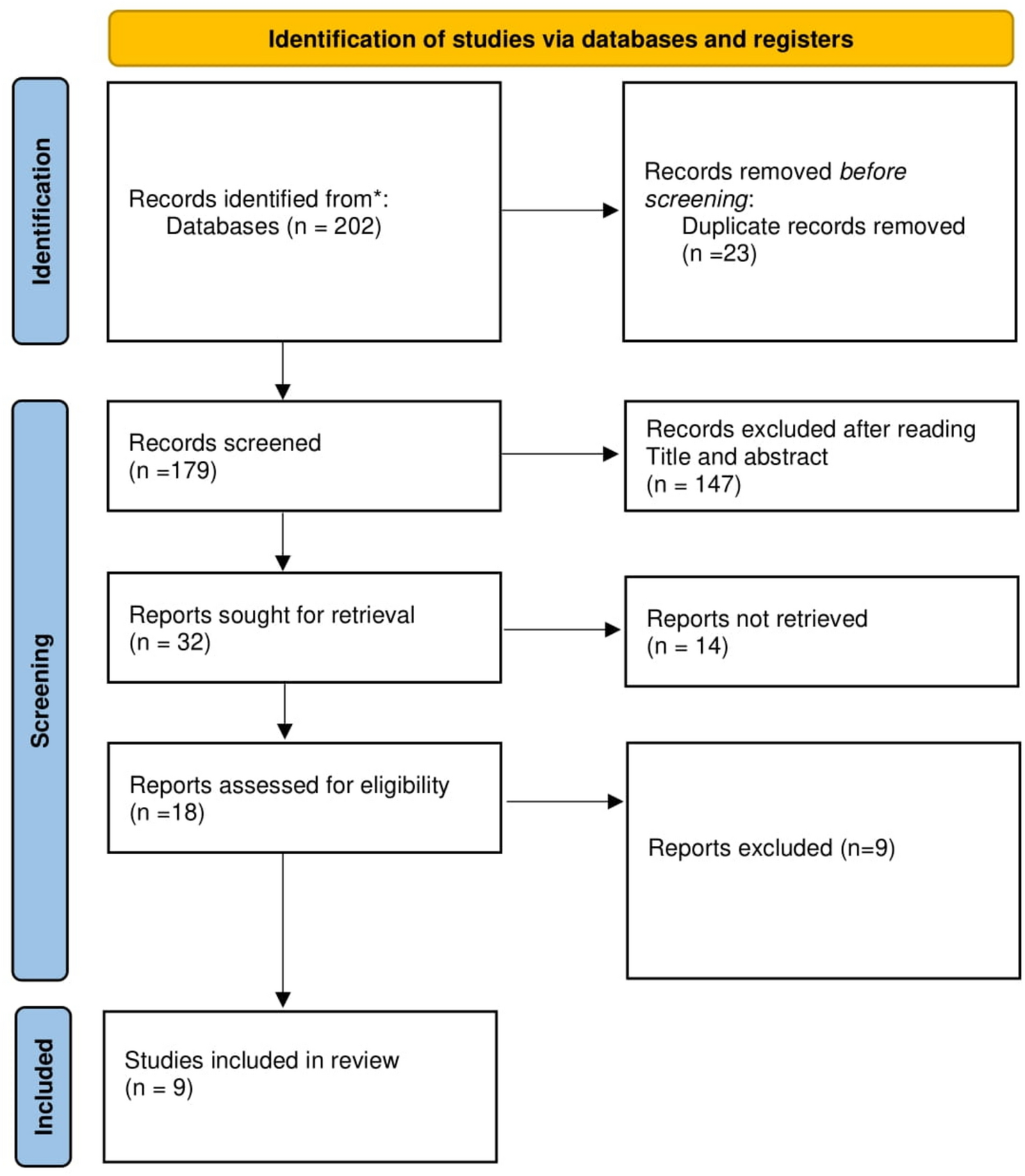
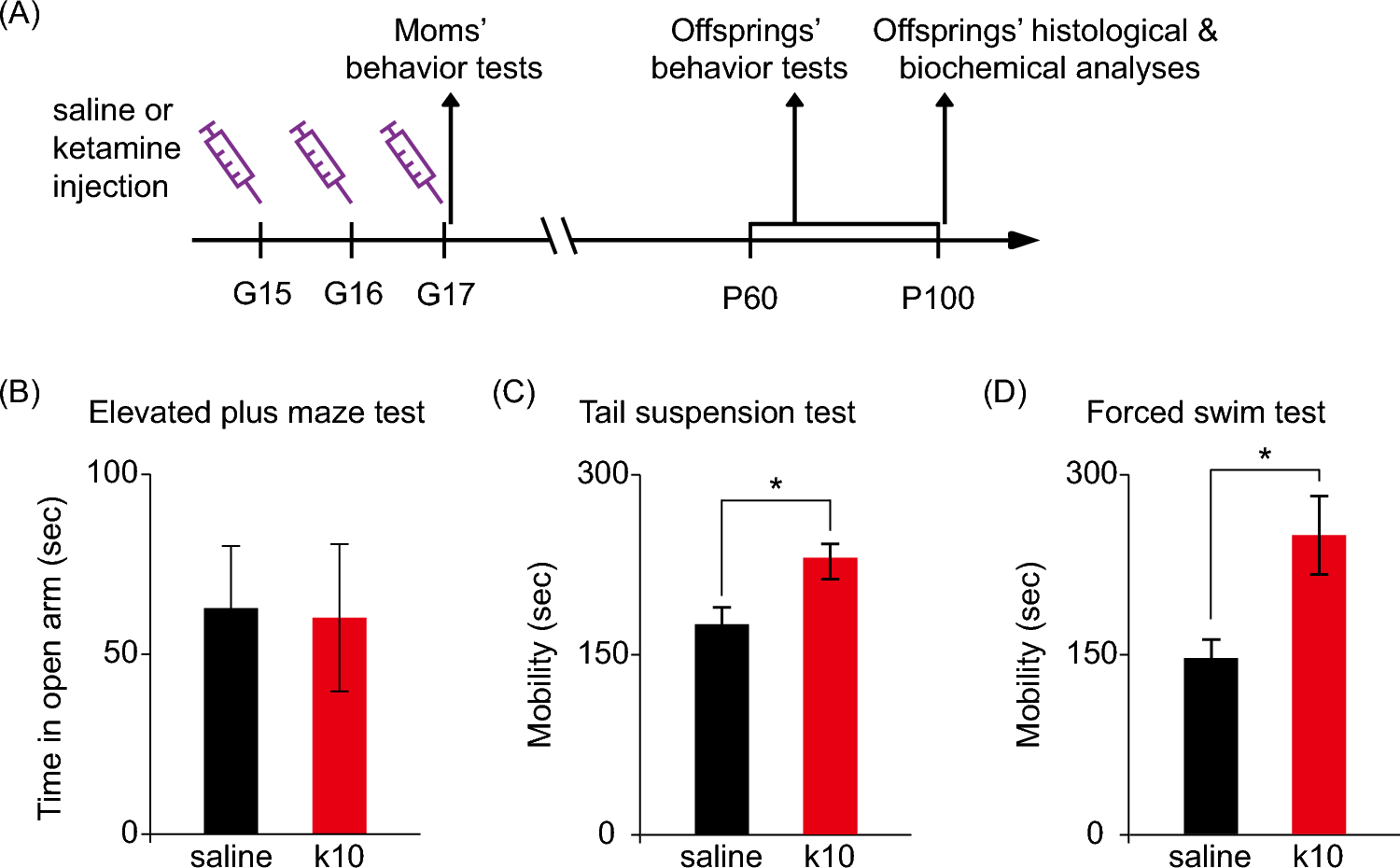
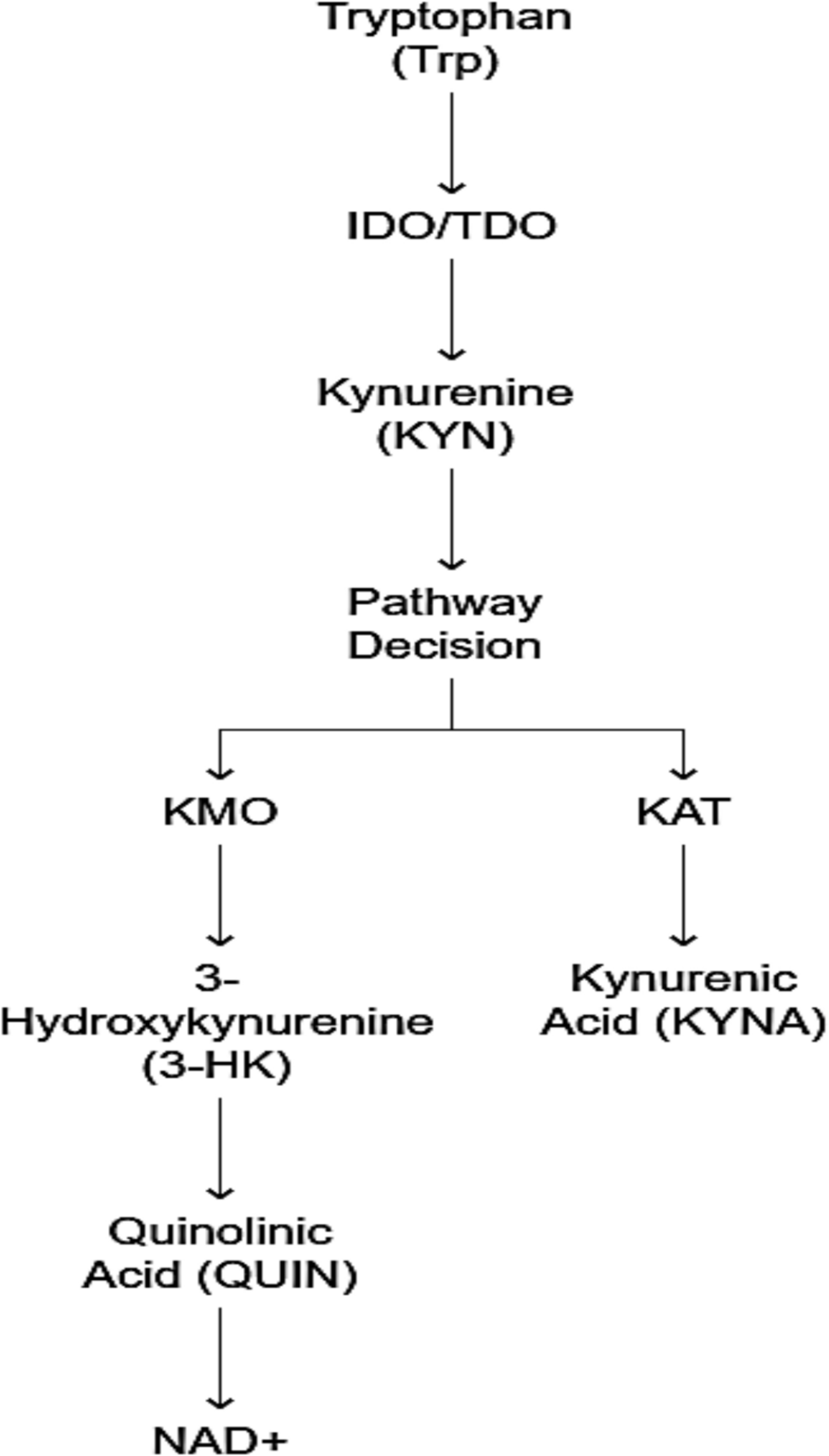
Remember me



Signaling molecules in NEI systems were collected using key words “neurotransmitters”, “neuropeptides”, “hormones”, and “cytokines” (detailed in “Materials and methods”). As a result, a total of 3144 protein-coding genes (2715 genes and 429 TFs) and 581 miRNA signaling molecules in NEI systems were obtained. Among them, there were 147 miRNAs, 578 genes, and 18 TFs in nervous system, 414 miRNAs, 1092 genes, and 259 TFs in endocrine system, and 435 miRNAs, 1473 genes, and 262 TFs in immune system (Supplementary Table S1). The Venn diagrams of the collected NEI miRNA, gene and TF signaling molecules were shown in Supplementary Fig. S1a, b, and c, most of the signaling molecules in nervous system were also the signaling molecules of the other two systems.
Background NEI-miRTF-NBackground NEI-miRTF-N was constructed by integrating NEI-related miRNA-TF FFLs, which were identified through NEI signaling molecules and the interactions between them. As a result, 370,885 FFLs were obtained, comprising 230,214 miRNA-type FFLs (62.07%), 92,248 TF-type FFLs (24.87%), and 48,423 composite FFLs (13.06%) (Fig. 2a, Supplementary Table S6). The number of nodes and links in these FFLs was represented in Table 1. Merging these FFLs resulted in the background NEI-miRTF-N, which consists of 2,658 nodes (494 miRNAs, 1991 genes, and 173 TFs) and 98,629 edges. For the background NEI-miRTF-N, there were 123 miRNAs, 160 genes and 4 TFs in nervous system; 379 miRNAs, 925 genes and 117 TFs signaling molecules in endocrine system; 394 miRNAs,1,263genes and 102 TFs signaling molecules in immune system (Fig. 2b, c, and d).
Fig. 2
Background NEI-miRTF-N. a The distribution of three types of FFLs in background NEI-miRTF-N. b The Venn diagram of NEI signaling miRNAs. c The Venn diagram of NEI signaling genes. d The Venn diagram of NEI signaling TFs. e Degree distribution of all nodes in background NEI-miRTF-N and degree distribution of miRNAs, genes, and TFs. f The number of NEI signaling miRNAs, genes, and TFs in background NEI-miRTF-N. g The regulatory relationships between NEI signaling molecules in background NEI-miRTF-N. h The significance of regulation relationships between signaling molecules compared with theoretical expected cases using hypergeometric test
Table 1 Summary of three types of NEI-related miRNA-TF FFLsThe node degree and its distribution in NEI-miRTF-N were investigated. As shown in Fig. 2e, we found that only a small portion of nodes highly connected with other nodes, while most nodes had relatively small degrees. This was consistent with the characteristics of biological networks. The average node degree of miRNAs, genes, and TFs was 150.84 (range 2–1005), 44.34 (range 2–256), and 199.25 (range 2–1663), respectively. Additionally, the node betweenness was examined (Supplementary Fig. S2), miRNAs and TFs have higher degrees and betweenness, indicating their greater importance in the network.
We further examined the distribution of neural, endocrine, and immune signaling molecules and their interactions in the network. As demonstrated in Fig. 2f and g (Supplementary Tables S7 and S8), the number of immune signaling molecules is the highest, while the number of neural signaling molecules is the lowest. At the same time, the mutual regulation between immune signaling molecules and endocrine signaling molecules is also the most. Additionally, we investigated the significance of regulation relationships between signaling molecules compared with that of theoretical expectation using hypergeometric test. As shown in Fig. 2h and Supplementary Table S8, we found that signaling molecules in neural system significantly regulate those in endocrine and immune system, and signaling molecules in endocrine system significantly regulate those in immune systems (p-value < 0.0001). This implied that the directions of signal transduction might mainly be from nervous system to the endocrine and immune systems, as well as from the endocrine to immune system.
Disease-Specific NEI-miRTF-NDisease-specific NEI-miRTF-N was constructed by mapping the known disease molecules and SDE molecules to the background NEI-miRTF-N. The method is applicable to all the diseases, provided that known disease-related molecules and SDE molecules can be obtained. Depression, AD, and DCM were used as case studies. The acquisition of SDE molecules for the three diseases is as follows.
For depression, AD, and DCM, 12, 5, and 4 mRNA expression datasets and 2, 2, and 2 miRNA expression datasets were retained after filtration (details see Materials and methods). Molecules with p-value < 0.05 were selected as SDE molecules. To ensure the reliability of the data, up- and down-regulation of SDE molecules in multiple datasets were required to be consistent. According to UpSet plots, it is required that SDE genes were differentially expressed in at least 2, 5, and 3 datasets for mRNA expression of depression, AD, and DCM, respectively (Fig. 3a, d, g and Supplementary Table S9). Due to the limited number of miRNA datasets, SDE miRNAs were taken as union sets, and it is required that the miRNAs in the intersection set had consistent up- and down-regulation in datasets. Finally, 62 SDE miRNAs, 720 SDE genes, and 96 SDE TFs in depression; 102 SDE miRNAs, 1085 SDE genes, and 72 SDE TFs in AD; 259 SDE miRNAs, 858 SDE genes, and 97 SDE TFs in DCM were obtained (Supplementary Table S4).
Fig. 3
a UpSet plots of SDE genes for depression. b Disease-specific NEI-miRTF-N for depression. c The distribution of three types of FFLs for depression. d UpSet plots of SDE genes for AD. e Disease-specific NEI-miRTF-N for AD. f The distribution of three types of FFLs for AD. g UpSet plots of SDE genes for DCM. h Disease-specific NEI-miRTF-N for DCM. i The distribution of three types of FFLs for DCM
We thus constructed depression, AD and DCM-specific NEI-miRTF-Ns, respectively (Fig. 3b, e, and h, Supplementary Table S6). There were 162 nodes and 669 edges, 62 nodes and 181 edges, 124 nodes and 652 edges in the three networks, respectively. For depression-specific NEI-miRTF-Ns, 492 FFLs were included, comprising of 288 miRNA-type FFLs (58.54%), 159 TF-type FFLs (32.32%), and 45 composite FFLs (9.15%) (Fig. 3c). For AD-specific NEI-miRTF-Ns, 104 FFLs were included, comprising of 92 miRNA-type FFLs (88.46%), 10 TF-type FFLs (9.62%), and 2 composite FFLs (1.92%) (Fig. 3f). For DCM-specific NEI-miRTF-Ns, 526 FFLs were included, comprising of 458 miRNA-type FFLs (87.07%), 49 TF-type FFLs (9.32%), and 19 composite FFLs (3.61%) (Fig. 3i).
Significantly Dysregulated NEI-Related miRNA-TF Regulatory Pathways (NEI-miRTF-Ps) in Depression, AD, and DCMIn this study, considering dysregulated extent of the known disease molecules and SDE molecules, we developed an algorithm to identify significantly dysregulated NEI-miRTF-Ps in diseases (details see “Materials and Methods”). As a result, 553, 11, and 290 significantly dysregulated NEI-miRTF-Ps in depression, AD, and DCM were identified, respectively (Supplementary Tables S10, 11, and 12). For dysregulated NEI-miRTF-Ps in depression, there are 17 miRNAs, 9 mRNAs, and 16 TFs in them, which included 5, 33, and 27 signaling molecules for N, E and I systems, respectively. The length of these pathways ranges from 4 to 16. For dysregulated NEI-miRTF-Ps in AD, there are 8 miRNAs, 2 mRNAs and 2 TFs in them, which included 8, 11, and 10 signaling molecules for N, E and I systems, respectively. The length of these pathways ranges from 3 to 5. For dysregulated NEI-miRTF-Ps in DCM, there are 25 miRNAs, 42 mRNAs, and 5 TFs in them, which included 13, 69, and 42 signaling molecules for N, E, and I systems, respectively. The length of these pathways ranges from 3 to 11.
The top 5 dysregulated NEI-miRTF-Ps in depression, AD and DCM were investigated, which were shown in Table 2 and Fig. 4a, b and c. Although the molecules in the pathway and the interactions between them were collected from relevant databases, most of them have not been experimentally validated. We examined these dysregulated NEI-miRTF-Ps from four aspects: (1) The associations between the molecules and diseases, as well as NEI systems. (2) The regulatory relationships between the molecules in the pathway. (3) The relations between biological functions of the molecules and diseases, as well as NEI systems. (4) The comparison of these dysregulated NEI-miRTF-Ps with the known disease-related KEGG pathways.
Table 2 The top 5 significantly dysregulated NEI-miRTF-Ps in depression, AD, and DCMFig. 4
a The top 5 significantly dysregulated NEI-miRTF-Ps in depression. b The top 5 significantly dysregulated NEI-miRTF-Ps in AD. c The top 5 significantly dysregulated NEI-miRTF-Ps in DCM. d Literature validation of significantly enriched biological functions of molecules from the top 5 dysregulated NEI-miRTF-Ps in depression. e Literature validation of significantly enriched biological functions of molecules from the top 5 dysregulated NEI-miRTF-Ps in AD. f Literature validation of significantly enriched biological functions of molecules from the top 5 dysregulated NEI-miRTF-Ps in DCM
We performed a comprehensive literature search, as represented in Supplementary Tables S13 and S14, the vast majority of molecules are associated with N, E or I and corresponding diseases, and most of the regulatory relationships between molecules have been verified. Additionally, for each pathway in top 5 significantly dysregulated NEI-miRTF-Ps, functional enrichment analysis was implemented using g:Profiler (Kolberg et al. 2023), significantly enriched GO BP terms and KEGG pathways were identified with Benjamini–Hochberg adjusted p-value < 0.05 (Supplementary Tables S15, S16, and S17). As shown in Fig. 4d, e, and f, we found that the large majority of significantly enriched biological functions have been validated to be associated with corresponding diseases as well as the nervous, endocrine, or immune systems.
Furthermore, we searched the KEGG pathways of the three diseases. There were 60, 384, and 103 genes in depression, AD, and DCM KEGG pathways, respectively. For each disease, we examined the enrichment of genes of significantly dysregulated NEI-miRTF-Ps in the KEGG pathway by applying hypergeometric test. When only considering genes and TFs in dysregulated NEI-miRTF-Ps, the p-values were not significant for the three diseases. While taking into account of miRNAs, they were all significantly enriched (p-value = 0.001 for depression, p-value = 4.77 × 10⁻5 for AD, and p-value = 0.001 for DCM). When we limited it to the top 5 dysregulated NEI-miRTF-Ps, it was not significantly enriched for depression (p-value = 0.130), marginally significantly enriched for AD (p-value = 0.069), and significantly enriched for DCM (p-value = 0.019). The above results showed that the identified dysregulated NEI-miRTF-Ps were credible.
Comments (0)