
Remember me





Spiral arterial remodeling (SPA-REM) is a common phenomenon in patients with PE and is characterized by a lower proportion of appropriately remodeled spiral arteries. This dysregulation is associated with impaired SMC loss within the arterial wall. In this study, we systematically reviewed articles published over the past five years regarding the mechanisms underlying smooth muscle cell regression disorders in the wall of the uterine spiral artery (refer to Fig. S2-S3 for a detailed analysis process). Our findings indicate that phenotype switching and apoptosis are the most recognized mechanisms of smooth muscle cell regression in this context, with a notable increase in the number of studies focusing on phenotype switching each year. By utilizing immunofluorescence and Tunnel staining, SMC apoptosis and migration induced by phenotypic conversion during spiral artery remodeling were further evaluated (Fig. 1B). According to the quantitative analysis, during the remodeling process of SPA vascular wall, the proportion of SMC undergoing phenotypic switching was higher than the proportion of SMC undergoing apoptosis (Fig. 1C).
Fig. 1
Smooth muscle cell phenotype switching: key mechanism in uterine spiral artery remodeling. A Systematic reviews indicate that SMC apoptosis and phenotypic transformation have garnered significant attention in the context of uterine spiral artery remodeling. Furthermore, the number of studies focusing on smooth muscle cell phenotypic transformation has shown a consistent annual increase. B, C In first trimester, the proportion of SPAs-SMC undergoing phenotype switching is higher than that of apoptotic SPAs-SMC (Each group included 15 participants, and two sections from different parts were randomly selected for each participant. In Fig. 1B1 and B2, green arrows point to SMC that have undergone phenotypic switching, yellow arrows point to SMC that have undergone apoptosis). D UMAP visualization of the human decidual cell atlas single-cell transcriptomics data colored by cell type, SMC were identified and categorized into contractile and synthetic phenotypes based on the expression of distinct marker genes. E Pseudo-time analysis for the transition of SMC from a contractile phenotype to a synthetic phenotype in the early pregnancy (The timeline progresses from left to right, with state 1 in the leftmost chart representing the earliest stage along the developmental axis, while state 3 represents a later stage in the progression). F Dynamic changes of SPAs-SMC during SPA-REM process. The statistical methods used in the figures are all adjusted p-values obtained by the nonparametric Wilcoxon rank-sum test and Bonferroni correction, data represent the mean ± SD, ns P >0.05, * P < 0.05, ** P < 0.01, *** P < 0.001. SPAs (Spiral artery), SPA-REM (Spiral artery remodeling), SMC (Smooth muscle cells), UMAP (Uniform Manifold Approximation and Projection), α-SMA (α-smooth muscle actin), CK7 (Cytokeratin 7)
In order to investigate the impact of SMC phenotype switching on spiral artery remodeling, a single-cell RNA-seq data analysis was performed on decidual tissue samples from early pregnancy (Fig. 1D). Then the analysis focused on cells identified as smooth muscle cells for cluster analysis was performed, the identification of single-cell transcriptome data clusters is detailed in Fig. S4 A-G. Pseudo-chronological clustering analysis indicated that SMC phenotype switching occurs alongside the primary direction of SMC differentiation, gradually replacing contractile phenotype smooth muscle cells over time (Fig. 1E). Together, these results revealed that SMC phenotype switching is the main fate of SMC during uterine SPA-REM (Fig. 1F).
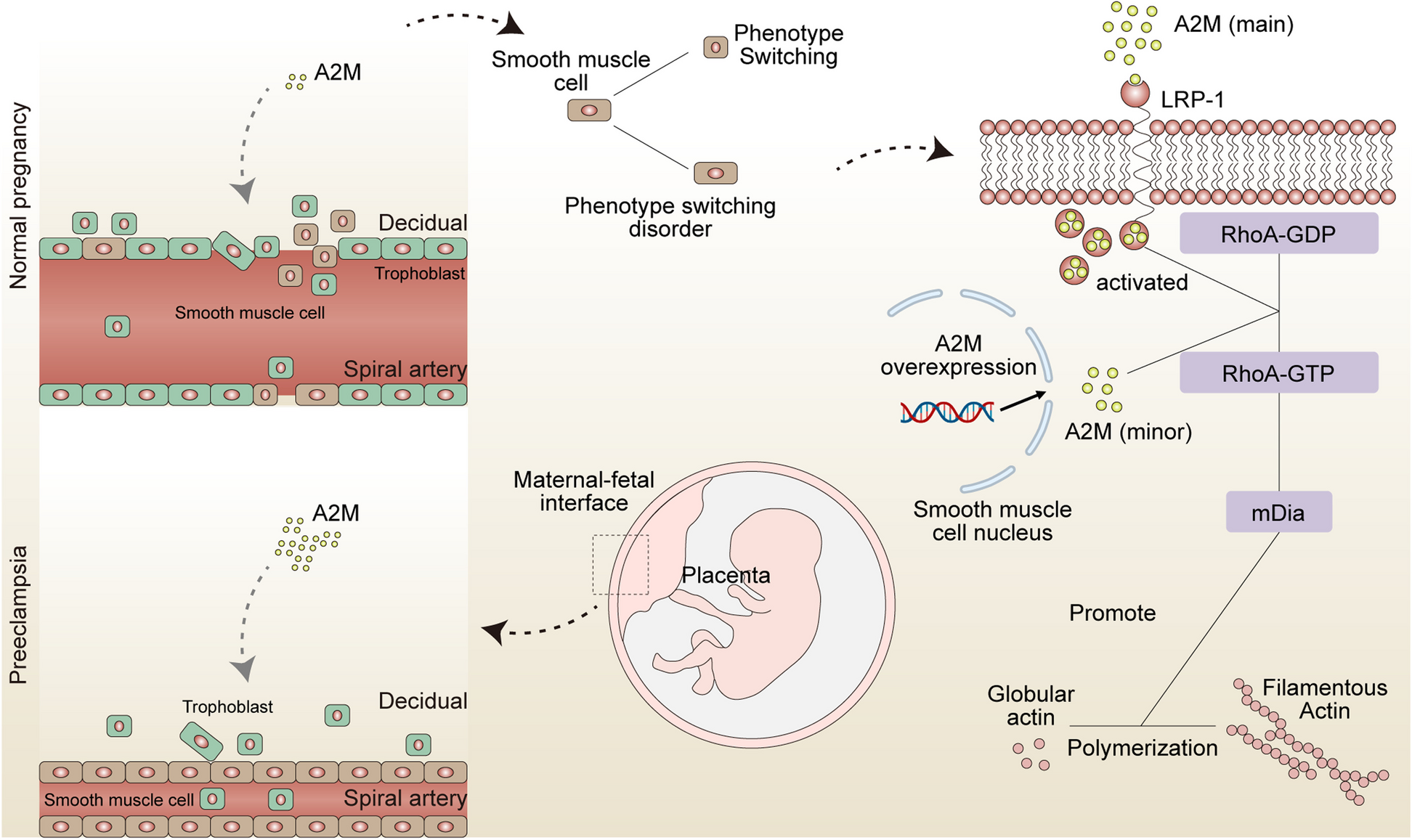
A2M is upregulated in smooth muscle cells of uterine spiral artery walls in patients with PE and its association with smooth muscle phenotype switching disorderIncreased A2M expression in decidual tissue and uterine spiral artery smooth muscles in patients with PE is positively correlated with the severity of spiral artery remodeling (Fig. 2A-D). Temporal analysis indicates a strong correlation between A2M expression and markers of smooth muscle contractile phenotype (Fig. 2E). Immunofluorescence staining of A2M and α-SMA confirms increased expression in vascular smooth muscle cells without phenotypic switching, additionally, fluorescence intensity analysis is consistent across different centrifugal distances (Fig. 2F-H), a significant positive correlation between fluorescence intensity of A2M and α-SMA was observed (R = 0.51, P < 0.05) (Fig. 2I).
Fig. 2
A2M is upregulated in smooth muscle cells of uterine spiral artery walls in patients with PE and Its association with smooth muscle phenotype switching disorder. A Representative images of A2M (red), α-SMA (green) and DAPI (blue) staining of uterine SPAs for normal pregnancy and (B) PE (The scale bar measures 175 μm, while the scale bar in the enlarged image on the right measures 75 μm. Each group consisted of 30 participants in the third trimester. For each participant, two morphologically intact decidual tissue sections from different regions were selected, and two random fields of view were chosen for quantitative analysis from each section). C Higher fluorescence intensity of A2M in the SMC of the decidual SPAs wall in pregnant women with PE. D Higher A2M fluorescence intensity in the smooth muscle of the SPAs wall with less remodeling. E Pseudo-time analysis showing consistent expression trends of A2M, α-SMA, and CNN1 in SMC. F Representative images of α-SMA (green), A2M (red) and DAPI (blue) staining of SMC at different levels of phenotype switching (The scale bar measures 90 μm, while the scale bar in the enlarged image below measures 40 μm. Each group consisted of 15 participants in the first trimester, and two sections from distinct regions were randomly selected for each participant. In the diagram, “1” indicates SMC that have undergone phenotype switching, while “2” denotes SMC that have retained their original phenotype without undergoing phenotype switching). G Unchanged of α-SMA and A2M fluorescence intensities with the cell centrifugal distance. H Higher fluorescence intensity of A2M in the SMC of the uterine SPAs with less phenotype switching (no displacement, spindle shape); I Pearson’s r correlations for fluorescence intensity of A2M and α-SMA in human decidual uterine SPAs. Data represent the mean ± SD, ns P >0.05, * P < 0.05, ** P < 0.01, *** P < 0.001. PE (PE), A2M (α2-macroglobulin), α-SMA (α-smooth muscle actin), CNN1 (Calponin 1), SMC (Smooth muscle cells), SPAs (Spiral artery)
A2M overexpression suppresses uterine spiral artery remodeling by hindering phenotypic switching of vascular smooth muscle cellsTo study the role of A2M in SPA-REM, A2M overexpression rats were studied, resulted in a phenotype similar to that of PE, which is characterized by impaired SPA-REM in both early and late pregnancy rats’ mesometrial triangle and superficial muscle layers (Fig. 3A-F). This indicates that A2M overexpression may play a role in inhibiting SPA-REM and contributing to the pathogenesis of PE.
Fig. 3
A2M overexpression suppresses rat spiral artery remodeling and smooth muscle cell phenotype switching. A-C Representative images of immunostained for α-SMA of placenta-decidua tissues of Ad-ctrl and Ad-A2M rats in early pregnancy. D-F Representative images of immunohistochemistry of placenta-decidua tissues of Ad-ctrl and Ad-A2M rats in early pregnancy (A, B, D, E) The scale bar is 300 μm, with the scale bar after enlargement being 50 μm, n=6. C, F Each group contained 6 rats, with 5 placenta-myometrium complexes randomly selected from each rat for morphological observation. Five fields of view were analyzed per complex, and results were presented as percentage stacked graphs). G Immunofluorescence staining of rat decidual and myometrium SPAs for α-SMA (red), CK7 (green) and DAPI (blue). H-J The fluorescence intensity of α-SMA in the decidua and superficial myometrium increased in the Ad-A2M group. I The decrease of the number of round smooth muscle cells in the decidua and superficial myometrium Ad-A2M group; J The increase of the number of spindle smooth muscle cells in the decidua and superficial myometrium in Ad-A2M group. Each group contained 6 rats, with 5 placenta-myometrium complexes randomly selected from each rat for morphological observation. Five fields of view were analyzed per complex. Each point in the statistical graph represents the average observation of a placenta-myometrium complex (The scale bar is 90 μm). K GO analysis of proteomic sequencing indicated that A2M overexpression may influence the phenotypic switching of smooth muscle cells by regulating processes such as cell morphology and actin filament organization. L Further KEGG analysis suggesting that A2M may affect actin filament organization through the actin cytoskeleton pathway. M Analysis of protein-protein interaction networks, N the MCC algorithm demonstrates the top 20 hub genes, with RhoA, CDC42, and Rac1 ranking highest. O Heatmap shows expression levels of genes in the networks. Significance levels indicated as * P < 0.05, ** P < 0.01, *** P < 0.001. Error bars represent the mean ± SD. A2M (α2-macroglobulin), α-SMA (α-smooth muscle actin), CK7 (Cytokeratin 7), GD (Gestational Day), MCC (Maximum Clique Centrality), GO (Gene Ontology), KEGG (Kyoto Encyclopedia of Genes and Genomes), RhoA (Ras Homolog Family Member A), mDia (Diaphanous-related formin 1), CDC42 (Cell Division Cycle 42)
Based on the observations in rat model decidual tissue, late-pregnancy rats with A2M overexpression exhibited more severe phenotypic switching disorders in spiral artery smooth muscle cells (Fig. 3G). Specifically, there was a higher α-SMA fluorescence intensity in the decidua and superficial muscle layer (Fig. 3H), as well as a greater proportion of impaired phenotype switching in the arterial wall helices. These cells displayed elongated spindle-shaped morphologies and a decrease in rounded or displaced cells when A2M was overexpressed (Fig. 3I, J).
Proteomic sequencing analysis was performed on rat mesometrial triangle and superficial uterine myometrium tissues. Gene Ontology (GO) analysis (Fig. 3K) identified ten significantly altered biological processes in the decidual tissues of rats with A2M overexpression, with a significant mechanism linked to smooth muscle cell phenotypic switching. Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis indicated that A2M overexpression may impact the cellular cytoskeleton through the ‘Regulation of cytoskeleton’ pathway (Fig. 3L). MCC analysis highlighted RhoA, Rac1, and CDC42 as the top three hub genes within the enriched pathways (Fig. 3M, N), with gene expression shown in Fig. 3O.
A2M overexpression suppresses smooth muscle cell phenotypic switching by upregulating the RhoA GTPase pathwayUpregulating A2M expression inhibited rat aortic smooth muscle cells (RASMC) phenotypic switching, resulting in reduced cell migration ability, increased contractile phenotype markers, and decreased synthetic phenotype markers (Fig. 4A-F). Conversely, A2M knockdown led to a higher degree of synthetic phenotype transition in RASMC.
Fig. 4
A2M is involved in the regulation of smooth muscle cell phenotype switching by promotes cytoskeleton polymerization. A Representative images of scratch assays to evaluate the effects of A2M on RASMC migration (Scale bar=100μm, repeat the experiment n=6); B A2M inhibited the migration ability of RASMC, and knockdown of A2M enhanced the migration ability of RASMC. C, D Western blot analysis demonstrating that A2M promoted the expression of contractile phenotype markers (α-SMA) and reduced synthetic marker expression (OPN). E Representative F-actin staining illustrated the effects of A2M on RASMC stress fibers (Scale bar=25μm). F Morphological observations indicating an increase in spindle cells and F-actin intensity with A2M overexpression. G-I Assessment of A2M effect on G- and F-actin fluorescence in RASMC (Scale bar=25μm, repeat the experiment n=6). A2M (α2-macroglobulin), RASMC (Rat thoracic aortic smooth muscle cell), OPN (Osteopontin), α-SMA (α-smooth muscle actin), RhoA (Ras Homolog Family Member A), mDia (Diaphanous-related formin 1)
F-actin and G-actin staining demonstrated that G-actin aggregation induced RASMC to contract and adopt a contractile phenotype. A2M overexpression promoted F-actin formation but inhibited G-actin depolymerization, resulting in increased F-actin fluorescence intensity and decreased G-actin fluorescence intensity. Conversely, downregulating A2M significantly increased G-actin fluorescence intensity while decreased F-actin fluorescence intensity. This reveals the role of A2M in maintaining the contractile phenotype by enhancing cytoskeletal aggregation (Fig. 4G-I).
In addition, we also explored the effect of A2M on RASMC apoptosis. In the rat decidual tissue, we observed that A2M overexpression reduced Caspase 3 expression surrounding spiral artery walls (Fig. S5A-B). Western blotting on rat decidual tissue indicated that A2M downregulated Caspase 3 while upregulated Bcl-2 expression (Fig. S5C-D). A2M overexpression inhibited RASMC apoptosis, while A2M knockdown increased apoptosis. PI assay demonstrated lower PI-positive cells with A2M overexpression and higher with A2M knockdown (Fig. S5E and F). Similar protein-level results can be found (Fig. S5G-H), with A2M overexpression increasing Bcl-2 expression and decreasing CASP3 expression. Flow cytometry showed more cells in the Q2 region with A2M overexpression and fewer with shA2M (Fig. S5I). On the other hand, these results were supported by the PCR (Fig. S5J).
A2M interacts directly with RhoA GTPase and promotes RhoA GTPase activationAccording to current knowledge, Rac1 and CDC42 are primarily involved in pseudopodia formation at the cell periphery, while RhoA mainly contributes to stress fiber formation in the cell center (Fig. 5A). F-actin fluorescence results suggested that A2M overexpression did not significantly increase pseudopodia formation (Fig. 5B and C), indicating that A2M may primarily regulate RhoA GTPase activity.
Fig. 5
A2M interacts directly with RhoA GTPase and promotes RhoA GTPase activation. A RhoA GTPase primarily regulates the formation of stress fibers and the lamellar cytoskeleton in SMC, whereas Rac1 and CDC42 are predominantly involved in promoting the formation of lamellipodia and filopodia cytoskeletal structures. B Representative immunofluorescence images of F-actin (red, the scale bars in the bove and below images are both 25μm, n=6). C Ad-A2M or Ad-shA2M did not affect the formation of pseudopodia in the edge of RASMC. D-E Representative immunoblots of A2M, RhoA, and mDia from human decidual and the quantified ratio of A2M/GAPDH, RhoA/GAPDH and mDia/GAPDH (n=30 for each group). H-I Representative immunoblots of RhoA, and mDia from RASMC, the quantified ratio of RhoA/GAPDH and mDia/GAPDH. J Representative images of A2M (red, the upper line), RhoA (red, the downlink), and DAPI (blue), (the scale bars are 25μm, with the scale bar after enlargement being 50μm, repeat the experiment n=6). K Molecular docking of A2M and RhoA. L Selected peptide hits of Rho GTPase family proteins associated with A2M were identified through mass spectrometry analysis. M-N Immunoprecipitation results showing that A2M and RhoA can bind to each other directly (repeat the experiment n=6). Error bars, mean ± SD, with significance levels indicated as * P < 0.05, ** P < 0.01, *** P < 0.001. A2M (α2-macroglobulin), RhoA (Ras Homolog Family Member A), mDia (Diaphanous-related formin 1), CDC42 (Cell Division Cycle 42), SMC (Smooth muscle cells), RASMC (Rat thoracic aortic smooth muscle cell)
Western blotting on human decidual tissue showed that A2M, RhoA, and mDia were upregulated and positively correlated in patients with PE (Fig. 5D-G). A2M overexpression increased RhoA and mDia protein levels, while A2M knockdown decreased their expression in smooth muscle cells (Fig. 5H, I). Further cell fluorescence experimental results suggest that A2M and RhoA are concentrated in the cytoplasm of RASMC (Fig. 5J).
To gain insight into the molecular mechanism between A2M and RhoA, protein-protein docking was performed (All LC-MS/MS analysis results are provided in Supplementary Material 2), and results suggest that A2M can stably bind to RhoA (Fig. 5K). To identify potential substrate proteins regulated by A2M, we used A2M immunoprecipitation coupled with mass spectrometry. Of the potential A2M binding proteins, RhoA piqued our interest as it has the highest sequest HT score (Fig. 5L). Further, the immunoprecipitation experiments result demonstrated binding between A2M and RhoA in RASMC, and direct binding could be observed between RhoA and A2M (Fig. 5M, N).
The decidual tissue secretions fail to rescue A2M overexpression-induced inhibition of smooth muscle cell phenotypic switchingTo investigate whether the effect of A2M on RASMC can be changed by the physiological environment of decidua, a Placenta Decidual Co-culture (PDC) model was established (Fig. 6A). Cytological experiments showed that decidual tissue secretions promoted smooth muscle cell migration and synthetic phenotypic marker expression, but did not reverse the effects of A2M on smooth muscle cells, resulting in inhibited migration and increased expression of smooth muscle contraction phenotype markers (Fig. 6B-E). Additionally, in a chorionic villus artery transplantation model, the PDC serum increased the proportion of synthetic phenotype smooth muscle cells while decreased spindle-shaped smooth muscle cells. However, this failed to rescue A2M overexpression-induced phenotypic switching (Fig. 6F, I).
Fig. 6
Placenta decidual co-culture medium fail to rescue A2M’s inhibition of smooth muscle cell phenotypic switching and apoptosis. A Schematic illustration of experiments. B, C Scratch assay evaluating RASMC migration (Scale bar=100μm, repeat the experiment n=6). D, E Representative immunoblots of α-SMA, OPN from human decidual and the quantified ratio of α-SMA/GAPDH, OPN/GAPDH (Repeat the experiment n=6). F Representative immunofluorescence staining of α-SMA (red), DAPI (blue) of chorionic villus artery transplantation model. G-I Quantitative analysis of the proportion of round SMC, displaced SMC, and fusiform SMC in the chorionic villus artery transplantation model. J, K PI staining assessing RASMC apoptosis rate (Scale bar=100μm, repeat the experiment n=6). L, M Representative immunoblot of RASMC, PDC medium up-regulated CASP3 and down-regulated Bcl-2 expression. N Representative images of Flow cytometry showing that PDC medium promoted RASMC apoptosis, whereas A2M+PDC medium reducing it (Repeat the experiment n=6). O Representative immunoblot of A2M in RASMC (Repeat the experiment n=6). Error bars, mean ± SD, with significance levels as * P < 0.05, ** P < 0.01, *** P < 0.001. PDC model (Placenta decidual co-culture model), A2M (α2-macroglobulin), RASMC (rat thoracic aortic smooth muscle cell), OPN (Osteopontin), α-SMA (α-smooth muscle actin), PI (Propidium Iodide Apoptosis Assay), CASP3 (Caspase-3), Bcl-2 (Bcl-2 apoptosis regulator)
In addition to focusing on the phenotype switching of RASMC, we also paid attention to the effect of apoptosis of RASMC. PDC serum could not counteract the inhibitory effect of A2M on apoptosis. Despite a significant increase in the proportion of PI-positive cells, decidual tissue failed to rescue the inhibitory effect of A2M on smooth muscle cell apoptosis in the PI assay (Fig. 6J-K). Western blotting confirmed that A2M promoted CASP 3 expression while inhibiting Bcl-2 expression at the protein level (Fig. 6L, M). Flow cytometry results showed an increase in cells in the Q2 zone with decidual tissue secretions, but this did not reverse the decrease observed in the A2M overexpression group. Overall, there was limited rescuing effect of decidual tissue secretions on endogenous A2M overexpression (Fig. 6N). Nevertheless, our data indicates that PDC serum do not exist an influence on smooth muscle’s A2M expression (Fig. 6O). In conclusion, these results indicate that, in a physiological environment, decidual tissue secretions cannot alter the inhibitory effects of A2M on the phenotypic transition and apoptosis of RASMC.
Blocking A2M/LRP1 interaction effectively alleviate smooth muscle cell phenotype switching disorderSingle-cell transcriptome analysis revealed co-localization of A2M and its receptor LRP-1 in macrophages and smooth muscle cells within decidual tissue (Fig. 7A-D). Current understanding suggests that the RAP primarily inhibits the A2M/LRP-1 interaction by competitively binding to LRP-1 (Fig. 7E). Further immunofluorescence experiments confirmed A2M and LRP-1 expression in smooth muscle cells of the uterine spiral artery wall. Comparison of LRP-1 fluorescence intensity in decidual tissue of preeclamptic and normal pregnant women showed no significant difference (Fig. 7F-I). Morphological observations of villous artery transplantation model demonstrated that RAP protein more effectively inhibited the contractile phenotype of SMC in the vessel wall compared to A2M gene knockdown (Fig. 7J, K). At the same time, the experimental results of qPCR (Fig. S6A), Western blotting (Fig. S6B-C) and flow cytometry (Fig. S6D) showed that RAP promoted the apoptosis of RASMC.
Fig. 7
Blocking A2M/LRP1 interaction effectively alleviate smooth muscle cell phenotype switching disorder. A, B Single-cell transcriptome analysis predicting the localization of A2M and its receptor LRP-1 in decidua. Violin plots depicted the gene expression profiles of A2M (C) and LRP-1 (D). E A schematic diagram for the competitive inhibition of A2M-LRP-1 by RAP. F, G Representative immunofluorescence staining of α-SMA (green), DAPI (blue) and A2M (red), with higher fluorescence intensity observed in PE. H, I Representative immunofluorescence staining of α-SMA (green), DAPI (blue) and LRP-1 (red) (F-I: Scale bar=100μm, the scale bar in the enlarged image is 35μm, Decidual tissues from 30 participants were selected, including 15 patients with PE and 15 normal pregnant women. Two complete tissue sections were randomly chosen from each participant, and two fields of view were randomly selected from each section for observation). J, K Representative images of villous vascular explants (Scale bar=100μm, the scale bar in the enlarged image is 90μm) showing that RAP promoted the presence of round smooth muscle cells and reduced spindle cells in the vascular wall. Six explant models were selected for each group, with three fields of view randomly selected from each model for statistical analysis, each point in the statistical graphs represented the average observation value of one explant model. Error bars, mean ± SD, with significance levels of * P < 0.05, ** P < 0.01, *** P < 0.001. LRP-1 (Low Density Lipoprotein Receptor-Related Protein 1), A2M (α2-macroglobulin), RAP (Receptor-associated protein), α-SMA (α-smooth muscle actin)
Receptor-associated proteins exacerbated SMC phenotype switching with reduce RhoA GTPase pathway activitySince knockdown A2M can reduce RhoA GTPase pathway activation, we sought to investigate the effect of RAP on it. Firstly, the results at the cytological level by PCR and Western blot experiments, revealing the inhibition of RAP on contractile phenotype markers and increased expression of the synthetic phenotype marker osteopontin (Fig. 8A-C). Then, the migration ability of RASMC in scratch experiments provide evidence to support the above conclusion (Fig. 8D, E).
Fig. 8
Receptor-associated proteins exacerbated SMC phenotype switching with reduce RhoA GTPase pathway activity. A The levels of α-SMA, CNN1, MYH11 and OPN were determined by qPCR. B-C Representative immunoblot of α-SMA and OPN on RASMC. D-E Scratch assay evaluating RASMC migration (Scale bar=100μm, repeat the experiment n=6). F Representative images of F-actin (red) and DAPI (blue) on RASMC (the scale bars are 25μm, with the scale bar after enlargement being 50μm, repeat the experiment n=6). G Quantification of mean optical density values of F-actin. H Proportion of round RASMC. I The proportion of RASMC with lamellipodia. J, K Representative immunoblot of RhoA and mDia on RASMC. L Schematic representation of the mechanism by which RAP blocks the binding of A2M to LRP-1 and modulates the activity of the RhoA GTPase pathway. A2M (α2-macroglobulin), α-SMA (α-smooth muscle actin), CNN1 (Calponin 1), MYH11 (Myosin Heavy Chain 11), OPN (Osteopontin), RhoA (Ras Homolog Family Member A), mDia (Diaphanous-related formin 1), CDC42 (Cell Division Cycle 42), RASMC (Rat thoracic aortic smooth muscle cell), RAP (Receptor-associated protein)
Cell fluorescence experiment results indicate that RAP can significantly reduce the fluorescence intensity of F-actin (Fig. 8F, G). In addition, we observed that the proportion of RASMC with round shape and fewer stress fibers increased in the RAP intervention group (Fig. 8H). In addition, RAP intervention did not increase the proportion of RASMC with pseudopodia (Fig. 8I). Furthermore, the experimental results suggest that RAP intervention significantly reduces the expression of mDia and RhoA at the protein level in RASMC (Fig. 8J, K). The above experimental results indicate that blocking A2M-LRP binding can significantly reduce the activity of the RhoA-GTPase pathway (Fig. 8L).
Receptor-associated proteins can rescue related disease phenotypes in pre-eclampsia ratsTo study the therapeutic potential of RAP for preeclampsia, we constructed LPS preeclampsia rat model, then injection of RAP via tail vein (Fig. 9A). According to the protein-protein interaction analysis, it is clear that RAP effectively binds to both A2M and LRP-1 (Fig. 9B). In the PE rat model induced by LPS, maternal blood pressure levels were reduced after tail vein injection of RAP, validating the initial hypothesis (Fig. 9C, D). Ultrasound evaluation of rat uterine artery pulsatility index and resistance index showed significant decreases with RAP treatment in LPS-induced preeclampsia (Fig. 9E-G). In statistical analysis, fetal and placental size/mass in the RAP-administered group is smaller compared to controls, along with reductions in fetal crown-rump length and placental diameter. Moreover, fewer offspring per litter were observed in the LPS group compared to controls. Although RAP administration increased placental size and weight of LPS-treated rats towards control levels, full restoration was not achieved (Fig. 9H, I). This suggests that RAP partially rescues the LPS-induced PE phenotype. Furthermore, RAP increased the number and proportion of remodeled spiral arteries in the superficial uterine myometrium, although slightly lower than controls (Fig. 9J-L).
Fig. 9
Receptor-associated Protein (RAP) Ameliorates the pre-eclampsia phenotype in LPS-induced pre-eclampsia model rat. A Schematic diagram of the experimental. B Molecular docking of RAP, A2M, and LRP1. C Systolic and (D) diastolic blood pressure of pregnant rat. E Ultrasound evaluation showing that the uterine artery pulsatility index (F) and resistance index (G) were increased in the LPS group, while there was no significant difference between the RAP+LPS group and the control group. H, I Representative images of fetal rat and uterus (The scale bar is 1 cm, points in the graph representing the average of all fetal and appendage measurements delivered by each rat). J Representative images of immunohistochemistry of uterine spiral artery remodeling (Scale bar=300μm, the scale bar in the enlarged image is 150μm, three placenta-myometrium complexes randomly selected from each rat in each group, and 3 fields of view randomly selected for each complex for statistical analysis. Each point represents the observation of one placenta-myometrium complex, and the average of the results was calculated). K, L In the LPS group, the number and remodeling rate of blood vessels in the decidua and superficial muscle layer were reduced, whereas in the RAP+LPS group, these values were similar to those in the control group. M, N Representative immunofluorescence staining of rat decidual spiral artery α-SMA (red) and CK7 (green) showing that the roundness and displacement SMC rate in the LPS group were reduced, and the spindle SMC rate were increased (Scale bar=100μm, the scale bar in the enlarged image is 40μm, three random fields of view of 5 placenta-myometrium complexes selected from 6 rats in each group for observation, with each point in the statistical graph representing a placenta-myometrium complex and reflecting the average of the observations). Error bars, mean ± SD, with significance levels of * P < 0.05, ** P < 0.01, *** P < 0.001. A2M (α2-macroglobulin), LRP-1 (Low Density Lipoprotein Receptor-Related Protein 1), RAP (Receptor-associated protein), CK7 (Cytokeratin 7), α-SMA (α-smooth muscle actin), SMC (Smooth muscle cells), LPS (Lipopolysaccharide)
As shown in morphological analysis images, RAP administration enhanced the inhibition of round cell proportion caused by LPS, although not fully restored to control levels. The proportion of displaced and spindle smooth muscle cells in spiral arteries of the superficial uterine myometrium in the RAP + LPS group is similar to that of controls (Fig. 9M, N). In conclusion, RAP administration partially mitigates the preeclampsia phenotype induced by LPS by influencing smooth muscle cell phenotypic switching and enhancing spiral artery remodeling.
Potential diagnostic and therapeutic implications of A2M for PEPlasma A2M levels were significantly higher in early-onset pre-eclampsia and preterm pre-eclampsia during the third trimester (Fig. S7A-D). These levels were found to be a reliable predictor of delivery timing in pregnant women (Fig. S7E).
A2M displayed potential to identify preterm birth across different gestational age categories, with increased expression correlating with longer gestational ages (Fig. S7F). Time-dependent ROC analysis highlighted the potential of A2M to differentiate between extremely preterm, very preterm, moderately preterm, and late preterm infants, with AUC values ranging from 0.692 to 0.799 (Fig. S7G). Finally, a diagnostic model combining pre-eclampsia with preterm birth was developed by incorporating A2M characteristics into a generalized linear model. The specific fitting formula was y=−42.864 + 0.134x, and the Akaike information criterion (AIC) was 72.416 to evaluate the model’s performance (Fig. S7H).
Comments (0)