
Remember me





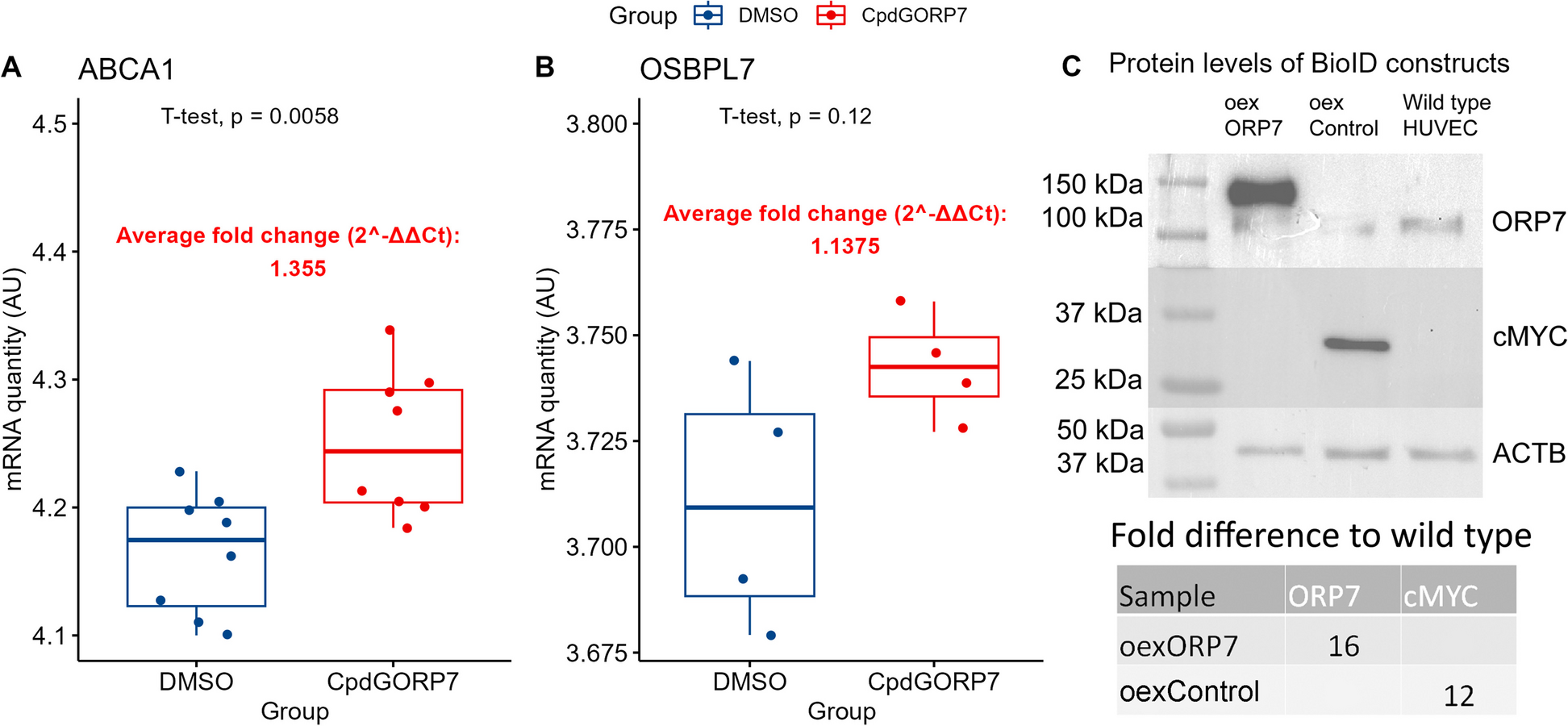

We initially cultured 4 established human melanoma cell lines – LM4, LM32, CHL-1 and A375M. We assessed each cell line for cell surface expression of CD44, EpCAM, CD24 and CD271 by flow cytometry (Fig. 1A). All cell lines were homogeneously CD44+EpCAM−, except for CHL-1 which contained a small number of CD44−EpCAM− cells. This demonstrates the mesenchymal character of all cell lines, in agreement with the ubiquitously high expression of the mesenchymal marker Vimentin in melanoma [12]. It also suggests that CD44, a CSC marker in epithelial cancers, does not discriminate sub-populations in melanoma. The surface expression of CD24 and CD271 was more variable between cell lines; the 4 lines expressed varying levels of CD271, whereas only CHL-1 expressed CD24. This gave rise to both CD24+CD271+ and CD24+CD271− sub-populations in the CHL-1 cell line only. Brightfield imaging (Fig. 1B) demonstrated a mesenchymal morphology for all cell lines, further underlining their mesenchymal character. However, there was heterogeneity between cell lines, with LM4 and LM32 having a homogenously mesenchymal morphology, whereas CHL-1 and A375M were more heterogeneous in their morphology, with clustering cobblestone regions suggesting an epithelial-like morphology. These cobblestone regions were more pronounced in CHL-1, which overall showed a greater level of morphological heterogeneity than the other three lines. In order to assess the CSC attributes of the four lines, we performed a tumoursphere assay. The tumoursphere assay tests the ability to survive and grow in suspension, and is a surrogate assay for CSC identity [21]; it enriches for cells that exhibit resistance to the oxidative stress and anoikis triggered by suspension culture, and continue to self-renew under these conditions [22]. We performed the tumoursphere assay under two conditions; in fully supplemented FAD medium with 10% serum, as used to maintain a proliferative undifferentiated phenotype in adherent culture (Fig. 1C, top), or in a defined serum-free medium that was developed for organoid culture and provokes differentiation (Fig. 1C, bottom) [23]. Interestingly, behaviour was starkly different under the two conditions; all four cell lines produced both primary and secondary spheres in the FAD medium, whereas only CHL-1 produced spheres in the more stringent serum-free conditions. Therefore, only the CHL-1 cell line retains a self-renewing CSC population under differentiation culture conditions.
Fig. 1
Cellular heterogeneity and stem cell attributes differ between melanoma cell lines. A Flow cytometric analysis of the four cell lines for CD44 and EpCAM (left) and CD24 and CD271 (right). The graphs show mean ± SEM of sub-populations in each of 4 quadrants based on ± gating set using isotype controls, as a percentage of the total cells, presented as stacked bars. Representative flow cytometry plots for each cell line are to the left of each graph. B Brightfield images of four melanoma cell lines in culture. C Sphere counts for the four cell lines, for primary spheres and secondary spheres after dissociation and re-plating, in serum-containing FAD medium (top) and serum-free medium (bottom). D Gene expression in FACS sorted CD24+ vs CD24− sub-populations. For each gene, expression was measured by QPCR and is represented as the expression level in CD24+ cells relative to that in CD24− cells. For each experiment, the number of biological repeats is indicated next to the graph. P-values were calculated using two-way ANOVA
Thus, of the four melanoma cell lines, only CHL-1 exhibits heterogeneous sub-populations based on any of the CSC markers derived from epithelial tumours. Both CD44 and CD24 were heterogeneously expressed in CHL-1, but only CD24 identified a minority positive sub-population that was absent from the other cell lines. CHL-1 was also the only cell line capable of producing tumourspheres under the more stringent serum-free conditions. We therefore focussed on CD24 as a potential marker of a sphere-forming CSC sub-population in CHL-1.
We performed QPCR analysis on FACS sorted CD24+ and CD24− sub-populations from CHL-1, for markers of the two recognised melanoma phenotypic states—melanocytic (proliferative) and neural crest-like (invasive) (Fig. 1D). Compared to CD24− cells, CD24+ cells had heightened expression of markers of the neural crest-like state (Zeb1 and Axl) and reduced expression of the melanocytic state marker MITF. The mesenchymal marker Vimentin was stably expressed across the two sub-populations. Interestingly, and contrary to the documented growth-arrested status of the neural crest-like state, the CD24+ sub-population had greatly increased Cyclin D expression. This suggested that CD24 may mark an alternative phenotypic state that has a dedifferentiated neural crest-like status whilst maintaining proliferative competence, as has been recently proposed [12]. The Notch signalling effectors Hes and Hey were reduced in the CD24+ sub-population, which is contrary to the documented role of Notch signalling in the CD271+ neural crest-like state [24, 25]. This is further evidence that CD24 may mark an alternative phenotypic state. Therefore, we focussed on CD24 alongside CD271 for further investigations in the CHL-1 cell line in order to determine whether this cell line contains a CSC sub-population that is absent from the other melanoma cell lines.
CD24 and CD271 mark an invasive stem cell sub-population in the CHL-1 melanoma cell lineWe FACS sorted four sub-populations (CD24+CD271+, CD24+CD271−, CD271+CD24− and CD24−CD271−) from the CHL-1 sub-line and performed a range of functional CSC assays. Using the double-negative CD24−CD271− sub-population as a baseline, we found that only the CD24+CD271+ sub-population produced a significantly increased number of tumourspheres (using the more stringent serum-free condition) (Fig. 2A). It also produced an increased number of colonies in a colony formation assay, another test for self-renewal (Fig. 2B). Therefore, the CD24+CD271+ sub-population exhibits enhanced anoikis resistance and self-renewal. To assess lineage plasticity, we cultured each sorted sub-population for 7 days and then re-assessed the sub-population distribution (Fig. 2C). Plasticity was much more pronounced in the CD24+CD271+ sub-population, which gave rise to large numbers of cells in each of the other three sub-populations. In contrast, the CD24+CD271− and CD24−CD271+ sub-populations gave rise mostly to CD24−CD271− cells, and the CD24−CD271− sub-population produced few cells in any of the other sub-populations. Therefore, the CSC attributes of self-renewal and lineage plasticity reside mainly in the CD24+CD271+ sub-population.
Fig. 2
CD24 and CD271 mark a stem cell sub-population in the CHL-1 melanoma line. A Primary sphere counts for four FACS sorted sub-populations from the 4 quadrants based on ± gating set using isotype controls. Results are presented as paired individual data points (due to considerable baseline variation between repeats), with each of the three marker-positive sub-populations compared individually to the CD24-CD271-sub-population. P-values were calculated using paired ANOVA (due to the variation between repeats). B Colony counts in 2D culture for the four FACS sorted sub-populations, with representative images of crystal violet stained colonies for each sub-population. P-values were calculated using unpaired ANOVA. C Flow cytometric analysis for CD24 and CD271, 7 days after sorting and re-plating the four sub-populations, with accompanying brightfield images of the four sorted sub-populations in culture. The graph shows mean ± SEM of sub-populations in each of 4 quadrants based on ± gating set using isotype controls, as a percentage of the total cells, presented as stacked bars. P-values were calculated using two-way ANOVA. Representative flow cytometry plots for each re-plated sub-population are to the left of the graph. For each experiment, the number of biological repeats is indicated next to the graph
We next assessed the ability of the four CHL-1 sub-populations to drive invasive dissemination. The CD24+CD271+ sub-population exhibited enhanced migration in a transwell migration assay (Fig. 3A). For a more stringent assessment of invasive ability, we developed a 3D matrigel-collagen invasion assay through adaptation of a previously published method [26]. In this assay, the melanoma cells are embedded in a matrigel ‘plug’ that is then itself embedded within a larger collagen outer gel. In this way, the degree of invasion of melanoma cells from the matrigel plug into the surrounding collagen can be assessed. The matrigel is rich in basement membrane components and thus provides an environment somewhat similar to the in situ tumour environment for a stringent assessment of invasive ability. Flattening the resulting 3D invasion into a 2D maximum intensity projection and plotting the average distance of each invading cell from the centre of the matrigel plug enabled assessment of the invasive ability of each sorted sub-population (Fig. 3B). Both the CD24+CD271+ and CD24+CD271− sub-populations had enhanced invasive ability compared to the CD24−CD271− sub-population.
Fig. 3
The CD24+CD271+ sub-population in the CHL-1 melanoma line has enhanced ability to migrate and invade. A The number of cells that have migrated to the other side of the membrane in transwell assays, for each of the four FACS sorted sub-populations. B Invasion of the four FACS sorted sub-populations into a collagen gel, when seeded in an inner matrigel gel. Left: Representative images, with each blue diamond representing a cell, projected onto x–y co-ordinates (z co-ordinates are flattened to zero). Units are pixel numbers in the x–y plane, and the orange star indicates the centre of the inner matrigel gel from which distance was calculated for each invading cell. Cells remaining in the inner gel are excluded from the visualisation. Top right: Average invasion distance for the four FACS sorted sub-populations. Bottom right: A representative brightfield image of cells breaking away from the inner gel and invading into the surrounding collagen. For each experiment, the number of biological repeats is indicated next to the graph. P-values were calculated using unpaired ANOVA
The CD24+CD271+sub-population exhibits enhanced therapeutic resistanceMelanoma exhibits broad chemotherapeutic resistance, and also quickly develops resistance to targeted therapies [27]. CSCs have been reported to exhibit enhanced therapeutic resistance in many tumour types, including melanoma, and drive the development of tumour drug resistance [7, 28, 29]. In particular, CD24+ CSCs in epithelial tumours exhibit the highest resistance to the chemotherapy drugs paclitaxel and cisplatin [7], although the tyrosine kinase inhibitor dasatinib has been reported to target CD24+ CSCs in epithelial tumours [30]. Therefore, we assessed the relative response of the four CHL-1 sub-populations to therapeutic challenge with these drugs (Fig. 4A). The CD24+CD271+ and CD24+CD271− sub-populations became enriched after treatment with dasatinib (contrary to previous findings in epithelial tumours). Conversely, enrichment in response to paclitaxel and cisplatin was exhibited by the CD24+CD271+ and CD24−CD271+ sub-populations. Therefore, the CD24+CD271− and CD24−CD271+ sub-populations exhibit differing resistance profiles, but these converge on the CD24+CD271+ sub-population which is enriched in response to all three drugs.
Fig. 4
The CD24+CD271+ sub-population is drug resistant. A Surviving cells in the CHL-1 melanoma line, after 3-day treatment with the indicated drugs and a 2-day recovery. B–E Surviving cells in the A375M and CHL-1 melanoma lines, after 3-day treatment with the indicated drugs and a 2-day recovery. The asterisk “*” denotes p < 0.05. In the brightfield images in E, blue arrows indicate cells with altered morphology. F Changes in distribution of sub-populations after generation of dasatinib-resistant CHL-1 (left) and A375M (right) melanoma lines. G Changes in distribution of sub-populations after TGFβ treatment for 6 days on CHL-1 (left) and A375M (right) melanoma lines. For all experiments, flow cytometric analysis for CD24 and CD271 is shown. The CD24/CD271 graphs show mean ± SEM of sub-populations in each of 4 quadrants based on ± gating set using isotype controls, as a percentage of the total cells, presented as stacked bars (A, F, G) or side-by-side bars (D). P-values were calculated using two-way ANOVA (A, F, G) or paired t-test (C, D). Representative flow cytometry plots for each condition are to the left of each graph. For each experiment, the number of biological repeats is indicated next to the graph
We next investigated whether the presence of CD24+ and CD24+CD271+ sub-populations led to enhanced overall survival of the CHL-1 cell line when treated with melanoma therapeutic compounds, by comparing to the A375M cell line which contains a CD271+ sub-population but does not contain CD24+ cells. We first assessed sub-population enrichment (Fig. 4B, D). The A375M cell line, which is BRAF mutant, exhibited an enrichment of the CD271+ sub-population in response to the chemotherapeutic agent Dacarbazine and the targeted drugs Vemurafenib (a BRAF inhibitor), Trametinib (a MEK inhibitor) and SCH772984 (an ERK1 inhibitor). The CHL-1 cell line, which is ERK1 mutant and BRAF wild type, was insensitive to BRAF inhibition. It showed an enrichment of both CD24+ and CD24+CD271+ sub-populations in response to Dacarbazine and Trametinib, and significant enrichment of only the CD24+ sub-population in response to SCH772884. Comparing overall resistance of the cell lines based on surviving cell number (Fig. 4C), we observed that CHL-1 was much more proliferative than A375M under all conditions (including control) but displayed enhanced overall survival compared to A375M in response to MEK inhibition and ERK1 inhibition and no enhancement in overall survival in response to Dacarbazine. Being BRAF wild type, it was insensitive to BRAF Inhibition. Enhanced survival of CHL-1 over A375M in response to targeted agents, to which CD24+ characteristics appear to confer resistance, alongside no enhancement in survival to a chemotherapeutic agent, to which CD271+ characteristics (shared with A375M) appear to confer resistance, further highlights the differing CD24+ and CD271+ resistance profiles, and their confluence on the CD24+CD271+ sub-population.
Brightfield images of the cell lines after treatment (Fig. 4E) further demonstrate the enhanced survival of CHL-1 in response to MEK inhibition and ERK1 inhibition. In concert with the cell survival data, they also show the relatively minor effect of ERK1 inhibition on the CHL-1 cell line; it continues to proliferate and display the same cellular morphology as in control conditions. Conversely, Dacarbazine and MEK inhibition induce altered morphology in the CHL-1 cell line, with the emergence of cells that exhibit a flat and spread cytoplasm (indicated by blue arrows). This effect is more pronounced with Dacarbazine, and correlates strongly with the emergence of a CD24+CD271+ sub-population (Fig. 4D). This morphological effect is not observed in the A375M line, which continues to display the same cellular morphology under all control and treatment conditions.
To investigate effects on longer-term development of resistance, we treated with dasatinib, a compound that in short-term experiments targeted the CD271+ sub-population but simultaneously elicited enrichment of the CD24+ sub-populations that are absent in the A375M cell line. Long-term dasatinib treatment resulted in dasatinib resistant CHL-1 (Fig. 4F, left). This caused a marked shift in the sub-population distribution, with a big increase in the CD24+CD271− sub-population and corresponding loss of the CD24−CD271+ and CD24−CD271− sub-populations. This underlines the role of CD24 in marking dasatinib resistant cells in this cell line. However, we were also able to create dasatinib resistant A375M using the same method, despite the absence of CD24 + sub-populations in this line. In A375M, the CD24−CD271+ sub-population was lost, resulting in an entirely CD24−CD271− cell line (Fig. 4F, right). This stands in contrast to the previously described enrichment of the CD271+ sub-population upon creation of Vemurafenib resistant A375M [19]. Therefore CD271+CD24− cells, whilst resistant to BRAF inhibitor Vemurafenib, are particularly sensitive to tyrosine kinase inhibitor dasatinib. Nevertheless, the fact that we were able to create a resistant cell line from A375M, which lacks CD24 + sub-populations, indicates that long-term resistance to CSC-targeted therapy can also be induced within the CD24−CD271− sub-population. This suggests that combination therapy, treating with CSC-targeted compounds alongside agents that kill the bulk CD24−CD271− cell population, may be required for successful melanoma therapy.
We next considered the potential role for an amoeboid phenotype in the enhanced invasive ability of the CD24+ sub-populations. Amoeboid cells are known to cluster at the tumour invasive front in melanoma and drive metastatic dissemination [31], and this phenotype is induced by TGFβ treatment [32]. We therefore treated the CHL-1 and A375M cell lines with TGFβ and observed the effect on the CD24 and CD271 marked sub-populations (Fig. 4G). In both lines, TGFβ caused an expansion of the CD24−CD271+ sub-population. In CHL-1, the CD24+CD271− sub-population was reduced and there was no change in the CD24+CD271+ sub-population. Therefore, TGFβ induces the CD24−CD271+ sub-population, indicating that an amoeboid phenotype is unlikely to be associated with the CD24+CD271+ CSC sub-population.
A CD24+CD271+sub-population in human melanoma tumour specimensWe investigated whether a CD24+CD271+ CSC sub-population exists in human primary melanoma specimens. Formalin fixed paraffin embedded (FFPE) sections of 31 melanomas were examined by a dermatopathologist, and included 19 superficial spreading, 11 nodular, and 1 acral lentiginous melanoma (Table 1). We developed a method for immunofluorescent co-staining for CD24 and CD271, combined with automated imaging and image stitching to create high-resolution images of entire pathological block sections. This enabled accurate assessment of CD24 and CD271 co-staining in each of the specimens, which was performed by both a researcher and a pathologist, guided by the matched H&E specimen to ensure that analysis of co-staining was restricted to melanoma cells (Fig. 5). Of the 31 tumours, 15 (48%) had evidence of CD24−CD271+ melanoma cells. These were specific to the invasive front in 5 (16%) tumours. Only 3 (10%) tumours had evidence of CD24+CD271+ melanoma cells and, interestingly, CD24+CD271− melanoma cells were also restricted to these same 3 tumours. In 2 of these tumours, the staining was specific to the invasive front. These findings confirm the existence of a CD24+CD271+ sub-population that localises to the invasive front in human melanoma but, in agreement with our panel of melanoma cell lines, is only present in a minority (3/31, 10%) of tumours.
Table 1 Pathological assessment of CD24 and CD271 staining in 31 melanoma specimensFig. 5
A CD24+CD271+ sub-population in human melanoma tumour specimens. A H&E of a human melanoma specimen, showing regions of melanoma cells (black arrows). B Isotype control staining. C Melanoma specimen with a region of CD24+CD271+ staining. Blue—DAPI nuclear stain; Green—CD24; Red—CD271. D Magnification of the region of CD24+CD271+ staining. Displayed images are image-stitches of multiple fields of view at 40 × magnification
The occurrence of metastatic spread was strongly correlated with Breslow thickness at presentation, as is well established in melanoma [33]. It was not possible to regress out Breslow thickness in order to test association with marker staining profile within this small specimen cohort. BRAF/NRAS status was available for 10 of the specimens, and this showed no correlation between BRAF/NRAS status and CD24/CD271 staining. One of the cases containing CD24+CD271+ cells was BRAF mutant, and one was NRAS mutant. The third was not tested. There were also CD24−CD271+ and CD24−CD271− cases that were BRAF and NRAS mutant. CD24+CD271+ staining is therefore not specific to tumours that are BRAF/NRAS wildtype, nor to BRAF/NRAS mutant tumours.
A CD24+CD271+sub-population in published human melanoma specimen scRNAseq datasetsInterrogation of the TCGA melanoma cohort (cBioPortal) supported the conclusions from our study, showing no correlation of CD24 expression with BRAF, NRAS, or ERK1 mutation or expression (though only 2% of cases are ERK1 mutant). The Broad Institute melanoma immunotherapy scRNAseq dataset [34, 35] had one tumour out of 14 containing CD24+ cells. This was in a bi-modal distribution, with only some of the cells being CD24+. The one tumour that expressed CD24 was also the only tumour with a large number of cells expressing CD271. In this tumour (Mel110), 13% of the total malignant cell population was CD24+CD271+ (Fig. 6).
Fig. 6
Scatter plot of CD24 and CD271 (NGFR) expression in scRNAseq of tumour Mel110 from the Broad Institute melanoma immunotherapy dataset [34, 35]. Axis scales are gene expression expressed as log2[1 + (TPM/10)], where TPM is transcripts per million. Colour coding represents our designation of cells into the 4 phenotypes based on the bimodal expression. These four designated phenotypes were used to perform the differential expression analysis presented in Additional file 1: Table S1
Another scRNAseq study [36, 37] (https://marine-lab.shinyapps.io/Human_melanoma_scRNASeq/) produced the same outcome for CD24: only one patient out of 20 had CD24+ cells, and CD24 was expressed in a bi-modal distribution. However, unlike in the Broad Institute dataset, the two tumours in this study that had significant CD271 expression did not overlap with the tumour containing CD24+ cells. Interestingly, and in line with our in vitro findings, these two CD271+ tumours did not contain any CD24+CD271+ cells. The one CD24+ tumour did however contain CD24+CD271+ cells, at 7% of the total malignant cell population. These CD24+CD271+ cells clustered together in the mesenchymal-like / neural crest-like malignant clusters. In this study, these were the clusters that exhibited drug resistance.
We performed differential expression analysis for the 4 sub-populations present in the Mel110 tumour [34, 35] (Additional file 1: Table S1). Consideration of all genes differentially expressed with p-value < 0.01 (or the top 200 most significant genes where more than 200 had p < 0.01) yielded lists of genes indicating the following biological differences:
1.The CD24+CD271+ sub-population had increased neuronal cell survival, migration, cytoskeletal reorganisation, and cell division genetic networks. This is similar to the CD24+CD271− sub-population. However, compared to the CD24+CD271− sub-population, the CD24+CD271+ sub-population lacked increased Sox2 and ALDH and had greater enhancement of neuronal cell survival and migration. This possibly suggests a neural crest stem cell phenotype. The CD24+CD271+ sub-population also, seemingly to a lesser degree, shared some key EMT pathways with the CD24−CD271+ sub-population. This supports the concept that there is a confluence of CD24+ and CD271+ characteristics in the CD24+CD271+ sub-population.
2.The CD24+CD271− sub-population had increased cell division and cytoskeletal reorganisation networks, and detoxification enzyme expression. The CSC markers Sox2 and ALDH were also increased. The CD24+CD271− sub-population had decreased Notch1 and other Notch signalling components compared to all other sub-populations, in agreement with findings from our cell line study.
3.The CD24−CD271+ sub-population had increased signatures for mitochondrial energy production, EMT, extracellular matrix component production, ribosomal biogenesis, transcription, RNA processing and protein folding.
There was no correlation of either CD24 or CD271 with BRAF or ERK1 expression in these cells. Interestingly, NRAS was upregulated in the CD24+CD271+ sub-population. This suggests a potential role for high NRAS expression, but not mutant NRAS, in the CD24+CD271+ sub-population. However, in our experiments, inhibition of the NRAS effector MEK caused CD24+CD271+ population induction in the CHL-1 cell line. Perhaps, as with the chemotherapeutic drug Dacarbazine, confluence of CD24+ and CD271+ attributes enables resistance to MEK inhibitors despite heightened NRAS expression.
These findings across two scRNAseq datasets agree with the findings from the cell line and human tumour analysis in our study. The overall proportion of CD24+CD271+ cells in the two tumours that contain them (one from each dataset) is similar to that in the CHL1 cell line. Also, in both our cell line and tumour analysis, some samples contained CD271+ without CD24+CD271+ cells, but CD24+ and CD24+CD271+ cells always co-existed; this is also true in these scRNAseq cases.
Comments (0)