
Remember me
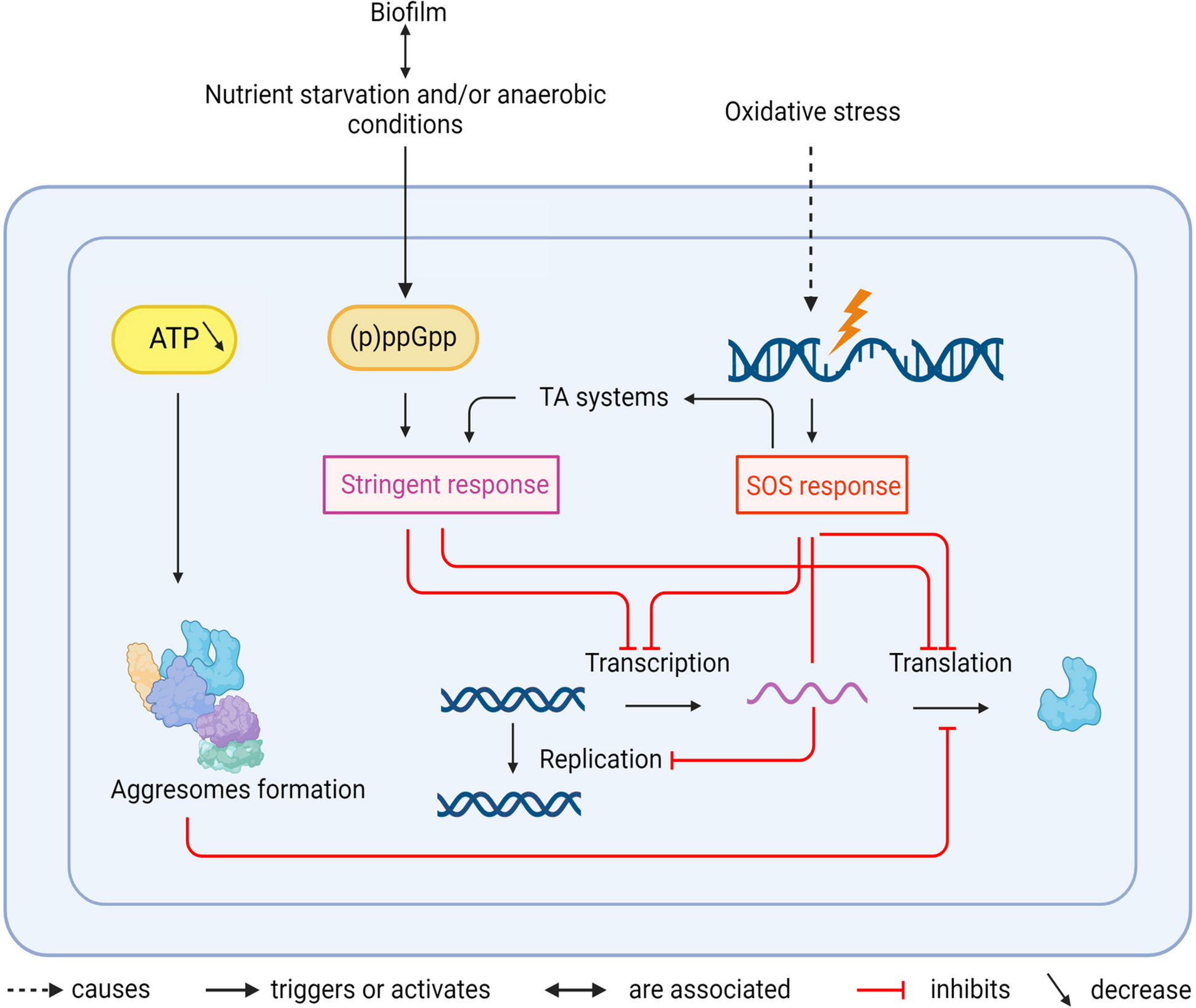

Recalcitrance is often linked to profound changes in cellular metabolism, which are listed below and in Fig. 1. Such changes can affect bacterial proliferation and make bacteria dormant. Dormancy is thus tightly linked to recalcitrance.
Fig. 1
Schematic summary of mechanisms leading to tolerance and persistence in a bacterial cell. Black arrows indicate activations, red arrows inhibitions. TA: Toxin/Antitoxin
Dormancy is induced by profound changes in cellular metabolismVarious pathways can lead to dormancy, all including a drop in cellular metabolism, such as a decrease in levels of ATP production, replication, transcription, or translation [13]. However, level of dormancy can differ within a bacterial population. Depending on the dormancy depth, time required for bacteria to resume growth can vary. When a bacterial population encounters favourable conditions for proliferation, some bacteria quickly regrow, while others take hours to days to resuscitate [12].
Dormancy is partly induced by depletion of intracellular ATP, which subsequently causes the formation of aggregates of endogenous proteins called aggresomes [12]. Proteins, mainly involved in replication, transcription and translation, aggregate and cannot function normally, blocking essential processes and inducing dormancy. The disintegration of aggresomes is a critical step for dormant bacteria to resume growth (a process also called resuscitation). Aggresomes disintegration is mediated by ATP replenishment, through the activity of proteases and chaperones. In E. coli, DnaK and ClpB proteins are recruited for resuscitation. DnaK binds to aggresomes, allowing the recruitment of the disaggregase ClpB [12].
A recent study reveals that tolerant and persistent cells enter different levels of dormancy [14]. Tolerance implies that a large majority of a bacterial population enters dormancy and thus survives to antibiotic exposure. These bacteria enter deep dormancy, stopping the translation of protein for transcription and repair of DNA. In contrast, persistent bacteria slow down their metabolism by reducing the production of ribosomal proteins and proteins involved in carbon metabolism and oxidative phosphorylation but keep a low level of transcription and translation [14].
(p)ppGpp and the stringent response play a major role in bacterial recalcitrance(p)ppGpp (guanosine penta/tetraphosphate) is a small molecule, which plays a predominant role during nutrient starvation, activating the so-called stringent response. In Gram-negative bacteria, (p)ppGpp is produced especially during starvation by proteins of the RelA/SpoT family. In Gram-positive bacteria, (p)ppGpp is controlled by Rel, which functions as both (p)ppGpp synthetase and hydrolase [15]. (p)ppGpp inhibits on one hand transcription and translation to slow down the metabolism of the bacteria. It also activates the expression of stress response genes and acts as a coordinator of many stress response mechanisms [15]. (p)ppGpp can also modulate the quorum sensing activity. In Pseudomonas aeruginosa, (p)ppGpp is required for the expression of the two quorum sensing systems RHL and LAS [16]. (p)ppGpp may thus indirectly affect formation of biofilm, which is an important player in bacterial tolerance (See below). (p)ppGpp and the stringent response have thus a key function for bacterial recalcitrance [15, 17].
Toxin-Antitoxin systems are responsible for persistence penetration in bacterial populationToxin-Antitoxin systems (TA systems) are composed of a toxin protein, which blocks a specific metabolic process, and of an antitoxin protein or RNA, which counteracts the toxin. Six different types of antitoxins have been described up to date, depending on the way they act to block the toxin (Reviewed in [18]). TA systems play an important role in persistence: an antimicrobial stress inactivates the antitoxins, allowing the toxins to block a metabolic process and therefore bacterial proliferation [8].
A well-known TA system is the HipAB system, which is widespread among Gram-negative species and was first described in E. coli [6]. The toxin HipA is a protein kinase that inhibits the glutamyl tRNA synthetase GltX by phosphorylating it. The inactivation of GltX leads to amino acid starvation and activation of the stringent response. The antitoxin HipB inactivates the toxin HipA directly by binding to it [19]. Upon a certain threshold of free HipA proteins, a subpopulation of cells enters dormancy and becomes persistent. The level of this threshold correlates with the level of penetration of persistence in the bacterial population. The heterogeneity in population is apparently caused by fluctuations of cellular quantity of toxin HipA, reaching or not the threshold. Mutations causing a decrease in the affinity of the toxin HipA for the antitoxin HipB, such as in the high-persistent mutant hipA7, can increase the percentage of population that enters dormancy [20].
DNA damage caused by oxidative stress can lead to recalcitranceROS stress can induce recalcitrance both directly and indirectlyReactive Oxygen Species (ROS) are produced by immune cells, macrophages in particular, in response to infection [21]. ROS such as superoxide (O2−) and hydroxyl radical (HO•) can damage DNA, lipids and proteins and cause the death of the bacterium [22]. However, these compounds can induce antibiotic tolerance, either through a direct activity on enzymes or indirectly via the SOS response [3]. They can directly inactivate aconitase, an enzyme of the Krebs cycle, as was shown in S. aureus infected macrophages [23]. In fact, ROS block the action of aconitase by attacking the Iron-Sulphur cluster in the active site of the enzyme. Consequently, the Krebs cycle is interrupted, bacteria synthesise low levels of ATP and their growth is slowed, inducing tolerance [23]. Beside ROS, nitric oxides can also induce recalcitrance through respiration and membrane potential inhibition [24, 25].
The SOS response is a mediator of dormancyThe SOS response is a bacterial process involved in the repair of damages to DNA made by ROS, among others [26]. SOS response causes a transient replication halt during which DNA can be repaired. It is regulated by two main actors: the repressor of the SOS regulon LexA and the sensor of DNA damage RecA. The repressor LexA is constitutively bound upstream of genes of the SOS regulon through a DNA sequence called the SOS box, thus repressing the expression of these genes when no DNA damages are detected. DNA damages induce accumulation of single stranded DNA that are recognized and bound by RecA. The formed nucleoprotein complex triggers the cleavage of LexA by autoproteolysis. SOS genes are thus expressed, including the sulA gene in E. coli, coding for a protein that stops cell division by sequestering the components of the cell division apparatus [27]. Furthermore, the SOS system includes a toxin-antitoxin system. IstR1, the antitoxin, is expressed at a basal level, whereas the toxin TisB is under LexA control [27]. When TisB concentration exceeds that of the antitoxin IstR1, free TisB forms pores in the bacterial inner membrane, decreasing the proton motive force and subsequently the activity of the ATP synthase [28]. Less ATP is produced, leading to dormancy of the bacterial cell.
Recently single bacteria RNAseq and CRISPR interference were used to identify common features of persister cells. This study suggests that Lon protease may be critical for persistence [29]. Lon is required to inactivate SulA to resume growth of recalcitrant bacteria. Deletion of either lon or sulA strongly decreases antibiotic survival [29].
A potential role for pili in toleranceBacterial aggregation and changes in motility can modulate antibiotic recalcitrance [30, 31]. Key factors of bacterial aggregation and motility are type IV pili [32]. Modifications of type IV pili have been recently proposed to affect tolerance in Neisseria gonorrhoeae and P. aeruginosa [31, 33]. Type IV pili are filamentous appendages composed of pilin proteins and involved in adhesion and motility, among others. PilE of N. gonorrhoeae undergoes antigenic variation, allowing immune escape. However, this has consequences on adhesion properties, aggregation and motility. It was observed that N. gonorrhoeae strains with stronger aggregation capabilities were also more tolerant to antimicrobial agents [33].
In contrast, non-motile P. aeruginosa strains lacking type IV PilD protein show increased antibiotic persistence to antibiotics. Such strains are commonly involved in chronic infections [31]. Taken together, these recent results indicate that type IV pili may play different roles in recalcitrance in different bacterial species.
Biofilm formation creates conditions conducive to persistenceBiofilms are the result of an aggregation of bacteria, generally through attachment to a substrate. These bacteria produce an extracellular matrix, composed of polysaccharides, proteins, extracellular DNA and other minor components [34]. This extracellular matrix acts as a very selective barrier, which makes the access to the bacteria more tricky for the antibiotics [35].
The persistence in biofilms can be the result of different mechanisms. In fact, biofilms create a microenvironment in which certain bacteria live in nutrient-poor, anaerobic conditions, especially at the core of the biofilm [35]. Bacteria in such an environment will have a low intracellular ATP level [36]. Consequently, starvation will trigger the expression of (p)ppGpp by the action of RelA/SpoT and therefore results in persistence [15].
Detection and study of recalcitrant bacteriaRecalcitrance is not trivial to study, because of its transient and heterogenous nature. It has been recently shown that CFU measurement to assess survival of bacteria following an antimicrobial treatment may be misleading for some antibiotics such as fluoroquinolones, since plating intoxicated bacteria on rich media may cause their death through reactivation of their metabolism [37]. Complementary techniques are thus useful to strengthen our knowledge of recalcitrance. Indeed, striking progress has been made in the detection and study of recalcitrant bacteria. The development of prokaryotic single-cell RNA sequencing has provided an essential tool to study heterogeneity in a bacterial population [38]. This was used recently to identify genes involved in persistence [29]. Other techniques such as FACS [39], microfluidics [40], microscopy [41] and nanomotion [42] have been proposed to increase our knowledge of recalcitrance. Taken together, these techniques will hopefully allow an easier detection of recalcitrant bacteria for both research and diagnosis purposes.
Persistence in vivo: a limited impact?Persistence has been clearly demonstrated in vitro and its in vivo importance has been repeatedly pinpointed in literature (Reviewed in [43]). However, recent study by Fanous et al. investigated the effect of a selection of antibiotics towards a mouse model of Salmonella infection [37]. The limited efficacy of the antibiotics could not be explained by stress-induced persistence. In contrast, nutrient depletion was causing slow growth, and subsequently increased population-level tolerance to antibiotics [37]. This indicates that further research is required to fully apprehend the role of persistence in vivo.
Strategies to eradicate recalcitrant bacteriaSince recalcitrant bacteria have a low metabolic activity, most antibiotics targeting active processes of bacterial cells, such as inhibitors of DNA, RNA, proteins, cell wall or folate synthesis, are unable to eradicate them. There are thus ongoing attempts to develop strategies to specifically target these recalcitrant bacteria. These strategies consist mainly in (i) the development of antimicrobial agents that are efficient towards recalcitrant bacteria and (ii) the development of drugs that modulate recalcitrance, as detailed below and in Fig. 2.
Fig. 2
Schematic summary of potential treatment strategies for recalcitrant bacteria
Antimicrobial agents efficient towards recalcitrant bacteriaTo be efficient towards recalcitrant bacteria, it is believed that antimicrobial agents should target processes that are independent of metabolic activity. Membrane active compounds are thus proposed as potential efficient anti-recalcitrant agents.
Membrane-active antimicrobial agentsMembrane disruptors directly target the structure, integrity, and function of the membrane [44]. Some antimicrobial peptides (AMPs) are membrane disruptors, making them potential anti-recalcitrance agents [45]. AMPs are short peptides, generally cationic, that are part of the innate immune defence of various organisms, including bacteria, plant, vertebrate and invertebrates [46]. Some AMPs that form pores in the membrane are effective on both metabolically active and dormant bacteria [47, 48]. This is for example the case for the AMP temporin G, which efficiently eradicates persistent S. aureus [47] and for LL-37 derived AMPs, which efficiently eradicate B. subtilis persisters, but not spores [39].
Polymyxins (Colistin and polymyxin B, among others) are polypeptidic antibiotics, used as reserve treatment towards infections caused by multi-resistant Gram-negative bacteria [49]. Polymyxins are positively charged polypeptides that bind to the outer membranes of Gram-negative bacteria via the negative charges of LPS [50]. They increase outer membrane permeability, pass into the periplasmic space and then disrupt the inner membrane, causing the bacteria to die. Colistin has been shown to be active against persistent P. aeruginosa in combination with other antibiotics [51].
Bacteriophages targeting dormant cellsBacteriophages, as natural predators of bacteria, can efficiently eradicate persistent bacteria. Bacteriophage-based treatment could thus be used as a complement to antibiotics to kill recalcitrant bacteria. This was recently applied on P. aeruginosa biofilm [52]. In contrast, in some cases, persistent bacteria can escape phages activity [53]. It is thus essential to identify the proper phages able to eradicate recalcitrant bacteria. Recently, Paride, a P. aeruginosa phage was shown to be efficient towards dormant bacteria both in vitro and in vivo, through a lytic cycle [54]. This indicates that some phages are able to bypass the low metabolic state of dormant bacteria, by a mechanism that has still to be investigated, in order to make them produce a new generation of phages.
Development of modulators of recalcitranceAn alternative strategy to directly target recalcitrant bacteria is to modulate either entry in or exit from recalcitrance. By forcing bacteria out of dormancy, or inhibiting dormancy initiation, one can avoid bacteria to be recalcitrant to antibiotics. Alternately, bacteria could be forced into an irreversible state of dormancy, avoiding the proliferation of recalcitrant bacteria after removal of the antibiotic.
Awakening dormant cells to make them sensitive to antibiotics againAwakening dormant cells could hit two birds with one stone: it may allow to ‘reset’ the metabolic processes targeted by the antibiotics, and to reactivate the energy-dependent influx pumps involved in antibiotic import. Metabolic stimulation with different kinds of sugars has been tested, with some success. Fumarate was identified as a potentiator of aminoglycoside towards tolerant P. aeruginosa in vitro [55]. This was also shown for tolerant E. coli, which could be sensitized to fluoroquinolones and other topoisomerase inhibitors through metabolic stimulation with several kinds of sugars [56]. Metabolization of sugars leads to ATP synthesis and reactivates proton motive force. A clinical trial (UROPOT) recently started, using mannitol as a metabolic stimulation in patients with asymptomatic E. coli and/or Klebsiella pneumoniae bacteriuria who undergo endourological surgeries [57]. This metabolic stimulation is aimed at eradicating bacteria tolerant to amikacin to avoid postsurgical complications.
Alternatively, antibiotic uptake can be activated through modification of pH gradient across bacterial membrane. Supplementation with L-arginine or L-glutamine, respectively, can modulate this pH gradient, thus enhancing aminoglycosides uptake [58, 59]. Effect of L-arginine in combination with tobramycin against P. aeruginosa biofilms was recently described, providing a potential treatment for recalcitrant P. aeruginosa in cystic fibrosis patients [60].
Inhibiting entry into dormancy(p)ppGpp is an important signalling molecule of the stringent response, involved in regulation of dormancy [61]. RelA, the protein responsible for (p)ppGpp biosynthesis is thus a potential target to fight recalcitrance. However, only few inhibitors of (p)ppGpp biosynthesis have been identified. Relacin was the first described (p)ppGpp biosynthesis inhibitor active towards Gram-positive bacteria, but its efficiency and pharmacokinetics properties were not compatible with a clinical usage [62]. Several attempts have been performed to find other RelA inhibitors, with limited success [63,64,65]. More recently, tetrone lactones, molecules structurally similar to vitamin C were shown to inhibit (p)ppGpp biosynthesis. They efficiently inhibit recalcitrance of Mycobacterium smegmatis, a close relative of M. tuberculosis [66].
Inhibition of TA systems activation is another strategy explored to fight recalcitrance. In this context, the Lon protease was identified as a promising target, since absence of Lon strongly decreased survival to antibiotics treatment [29]. Lon inhibitors have thus been tested to inhibit recalcitrance [67]. Bortezomib is an example of Lon inhibitor, which may be used in the future as an anti-recalcitrance agent [68].
Inhibition of polyphosphate kinase was also highlighted as a potential strategy to fight recalcitrance. Inorganic polyphosphate is required for persistence of A. baumannii, among others [69]. It is produced by polyphosphate kinase (PPK), which was shown to be inhibited by scutellarein. Scutellarein, a naturally occurring flavonoid, lowers recalcitrance of A. baumannii both in vitro and in vivo [70].
By pursuing the investigation of recalcitrance mechanisms, novel drug targets will most probably be described in the future, opening new paths for the development of anti-recalcitrance compounds. It has however to be mentioned that dormancy inhibitors may have a limited clinical benefit, since they should be applied at the early onset of infection and could be inefficient on already installed chronic infections.
Making dormancy irreversibleRecent advances have been made in the understanding of mechanisms involved in dormancy. The description of aggresomes formation, along with the description of different levels of dormancy opens the door to alternative strategies aimed at locking bacteria in an irreversible state of dormancy. This could be performed by inhibiting the disaggregation of proteins involved in resuscitation. Inhibitors of DnaK or ClpB could be developed for this purpose as recently proposed by Pu and colleagues [12, 71].
Targeting the communication between bacteria to prevent biofilm formationQuorum sensing is a major mediator of the communication between bacteria and plays a significant role in the biofilm formation. Therefore, it is a potential target for anti-persister treatments. Extensive ongoing research is performed in the field with numerous new potential quorum sensing inhibitors described recently, as reviewed elsewhere [72, 73]. However, quorum sensing inhibitors efficiency might be limited and have a rather narrow spectrum of action, since different bacterial species produce different quorum sensing molecules [74].
Comments (0)