
Remember me
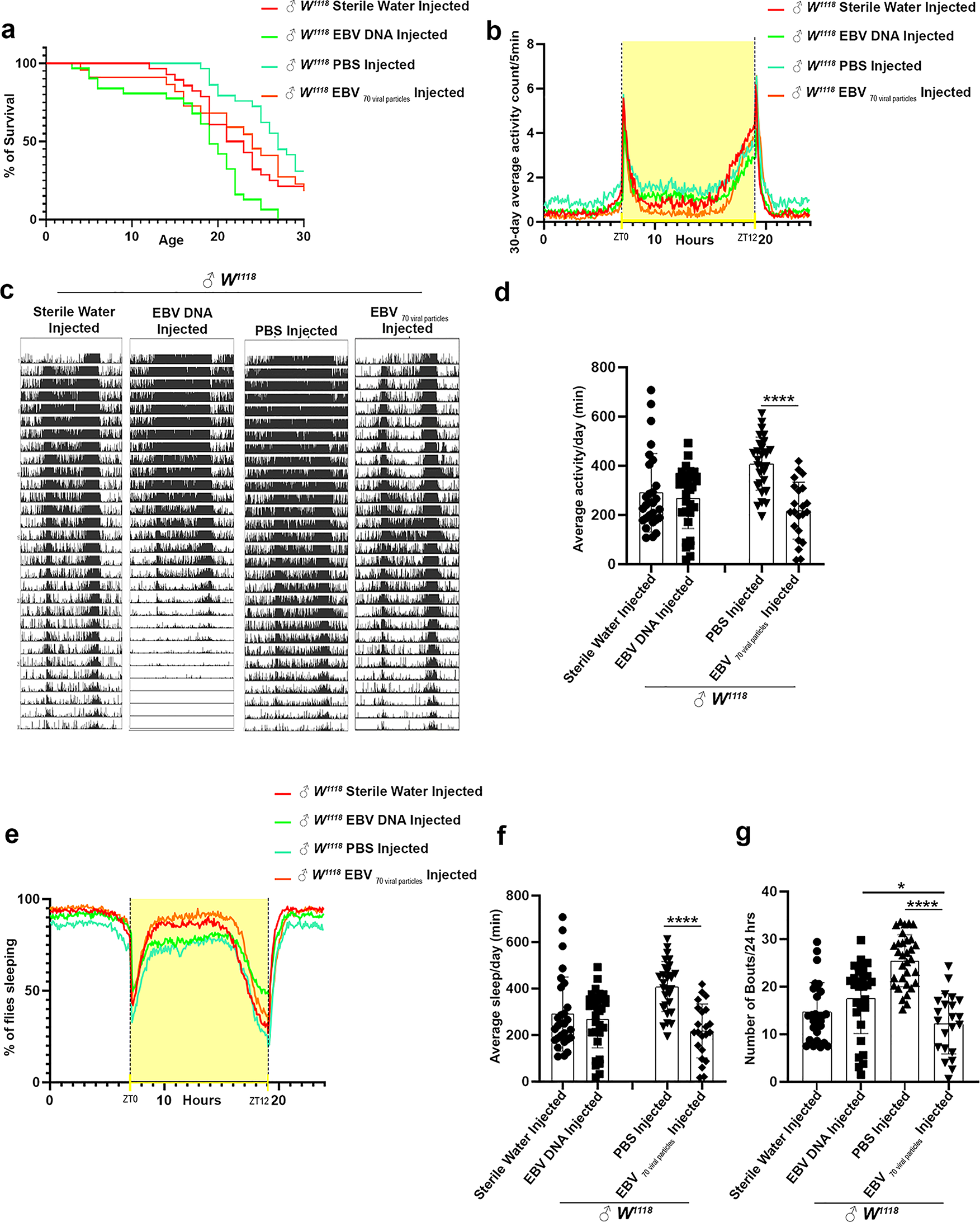

Flies injected with EBV DNA exhibited a significant decrease in lifespan, with a median survival of 19 days compared to 22 days for sterile water-injected flies (p = 0.0235) and 24 days for EBV viral particle-injected flies (p = 0.0242). Although flies injected with EBV viral particles also showed a reduced lifespan (median survival = 24 days), the difference was not statistically significant when compared to PBS-injected controls (median survival = 27 days, p = 0.0618) (Fig. 1a). These findings suggest that EBV DNA has a more pronounced effect on reducing lifespan than EBV viral particles.
Fig. 1
Lifespan, activity, and sleep plots of male W1118 injected either with EBV DNA or with 70 EBV particles. (a) Kaplan-Meier survival curve of male W1118 injected either with Sterile Water (n = 28), EBV DNA (n = 31), PBS (n = 32) or 70 EBV viral particles (n = 22). Outliers from each group were determined, and statistical significance was analyzed by the Mantel- Haenszel test. ns: p > 0.05; * p < 0.05; ** p < 0.005; *** p < 0.0005; **** p < 0.0001. (b) Representative plotted average fly activity profiles of males over 30 days. For each group, the locomotor activity levels of male flies were measured in 5-minute bins and then averaged to obtain a representative 24-hour activity profile. (c) Activity profile graph representing the average activity over 30 days, measured every 5 min. Drosophila melanogaster generally exhibits two activity bouts: one centered on ZT0 (morning peak) and the second around ZT12 (evening peak). (d) Average activity of the male flies over 24 h intervals using Kruskal-Wallis. (e) Sleep profile graph representing the percentage of flies engaged in sleep measured every 5 min over a 24 h period. ZT0 indicates the morning peak and ZT12 the evening peak. (f) Average sleep (minutes) of the flies over 24 h intervals. (g) Number of sleep bouts calculated during 24 h of spontaneous sleep. Kruskal-Wallis was used to determine the significance between groups. (* p < 0.05; ** p < 0.005; *** p < 0.0005; **** p < 0.0001. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 31, control PBS injected W1118: ♂n = 32; EBV 70 viral particles injected W1118: ♂n = 22.). Error bars represent standard deviation
Under light/dark conditions, Drosophila typically display a bimodal activity pattern, with peaks at the morning transition (ZT0) and the evening transition (ZT12), separated by a midday rest period or “siesta.” Circadian analysis revealed that both EBV DNA and EBV viral particle injections resulted in diminished locomotor activity, particularly during the anticipation phases before light-to-dark and dark-to-light transitions. Flies injected with EBV viral particles showed the lowest activity levels at ZT0, while those injected with EBV DNA exhibited a noticeable decline in activity before ZT12 (Fig. 1b, c). Overall, the mean daily activity was significantly reduced in both experimental groups compared to controls, with the most pronounced decrease observed in EBV viral particle-injected flies compared to PBS-injected controls (mean = 217.477 vs. 408.555, p < 0.0001) (Fig. 1d).
Alongside locomotor disruptions, sleep behaviors were also disrupted. Flies injected with EBV viral particles exhibited a significant increase in total daily sleep duration compared to PBS-injected flies (mean = 1236.59 min vs. 1087.16 min, p < 0.0001), whereas no significant differences were observed between EBV DNA-injected and sterile water-injected flies (Fig. 1e,f). The increased sleep observed in EBV viral particle-injected flies was accompanied by a decrease in sleep fragmentation, indicating by a reduced number of sleep bouts per 24-hour period (mean = 12.22 bouts) compared to PBS-injected flies (mean = 25.45 bouts, p < 0.0001). In contrast, EBV DNA-injected flies did not show significant differences in sleep fragmentation compared to sterile water-injected controls (Fig. 1g).
Day-night behavioral disruptions in EBV DNA and EBV viral injected fliesEngaging in day-night analysis provides a comprehensive examination of the behavioral variations exhibited by these fruit flies, offering invaluable insights into their sleep-wake cycles and responses to environmental cues throughout the day [28]. EBV viral particle injections resulted in a significant reduction in mean daily activity, with the effects being more pronounced in the EBV viral particle group during the day. While no significant differences were observed between EBV DNA and sterile water-injected flies (mean = 195 vs. 221 min during the day; mean = 73 vs. 96 min during the night), EBV viral particle-injected flies showed a sharp decrease in activity compared to PBS-injected flies (mean = 143.177 vs. 257.031 min, p = 0.0016 during the day; mean = 145 vs. 74.3 min, p = 0.0398 during the night) (Fig. 2a).
Fig. 2
Day-night dynamics of EBV DNA and EBV particles injected in Drosophila melanogaster. Average locomotor activity of the male flies over 12-hour intervals (day: light on, night: light off). (b) Average of daily sleep minutes for all flies over 12 h intervals over 30 days. (c) Number of sleep bouts was calculated during the day (lights on) and night-time (lights off). Statistical analysis performed using Kruskal-Wallis * p < 0.05; ** p < 0.005; *** p < 0.0005; **** p < 0.0001. (Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 31; control PBS injected W1118: ♂n = 32; EBV 70 viral particles injected W1118: ♂n = 22)
Sleep patterns differed between the two groups with an increase in sleep counts per min for EBV viral particles compared to EBV DNA (p value = 0.049 and p value < 0.0001 during day and night, respectively). EBV DNA-injected flies exhibited a significant reduction in sleep during both the day and night compared to sterile water-injected flies (p value = 0.0428 and p value < 0.0001 during day and night, respectively), whereas EBV viral particle-injected flies showed a significant increase in sleep duration in comparison with PBS control (p = 0.027) (Fig. 2b). Rather than inadequate sampling, the observed variability is most likely the result of biological variation among individual flies. This threshold is exceeded by the roughly 30 flies per condition in our study, which offers a trustworthy dataset for examining patterns of activity and sleep. The number of sleep bouts was also higher in EBV DNA-injected flies compared to EBV viral particle-injected flies during the day (p = 0.00493). However, no significant differences were observed between EBV DNA and sterile water-injected flies in terms of sleep fragmentation. In contrast, EBV viral particle-injected flies exhibited significantly fewer sleep bouts compared to PBS-injected flies during both day (mean = 7.58485 vs. 15.5729, p < 0.0001) and night (mean = 5.41212 vs. 10.8167, p = 0.0002), suggesting a more consolidated sleep pattern (Fig. 2c). EBV DNA and EBV viral particles induce distinct yet overlapping disruptions in lifespan, locomotor activity, and sleep patterns in Drosophila. While EBV DNA significantly shortens lifespan and reduces activity, its effects on sleep are minimal. In contrast, EBV viral particles lead to pronounced locomotor suppression and a substantial increase in total sleep duration and are accompanied by improved sleep consolidation. These findings highlight the differential impacts of EBV DNA and viral particles on behavioral and physiological functions, warranting further investigation into their underlying mechanisms.
Age-dependent behavioral changes in EBV DNA and EBV viral particle-injected fliesAnalyzing both young and old flies using the DAM is pivotal in uncovering age-related behavioral changes and patterns, shedding light on the dynamic interplay between aging and biological clock mechanisms; ‘young’ refers to 1–13 days old, and ‘old’ refers to 14–30 days old flies. The daily locomotor activity conducted in young versus old flies revealed remarkably different paradigms. EBV viral particle-injected young flies were notably less active than PBS-injected controls (mean = 303.126 min vs. 637.5 min, p = 0.0019) (Fig. 3a). These results suggest that EBV viral particles lead to a marked suppression of movement in young flies.
Fig. 3
Comparative behavioral analysis between young and old EBV DNA or EBV particles injected Drosophila melanogaster. (a) Average activity of the male flies over 24 h intervals. (b) Average sleep (minutes) of the male flies over 24 h intervals. (c) Number of sleep bouts was calculated during 24 h of spontaneous sleep. The flies are subdivided into two groups in an age-dependent manner since locomotion is age-dependent: ‘young’ refers to 1–13 days old, and ‘old’ refers to 14–30 days old. Kruskal-Wallis was used * p < 0.05; ** p < 0.005; *** p < 0.0005; **** p < 0.0001. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 31, control PBS injected W1118: ♂n = 32; EBV 70 viral particles injected W1118: ♂n = 22)
Sleep patterns varied significantly between young and old flies across injected groups. EBV DNA-injected flies exhibited a substantially reduced sleep duration compared to EBV viral particle-injected flies in both young and old generations (p = 0.0002 and p < 0.0001 respectively). Specifically, in old EBV DNA-injected flies, sleep duration was significantly lower than in sterile water-injected controls (mean = 1045.57 min vs. 1297.09 min, p < 0.0001) (Fig. 3b). Further differences emerged in sleep fragmentation. Young EBV DNA-injected flies exhibited a significantly higher number of sleep bouts compared to EBV viral particle-injected flies (p < 0.0001). In contrast, EBV viral particle-injected flies showed fewer sleep bouts compared to PBS-injected controls in both young (p < 0.0001) and old (p = 0.0013) generations (Fig. 3c). Age plays a crucial role in how EBV DNA and EBV viral particles affect locomotor activity and sleep in Drosophila. While young EBV DNA-injected flies exhibit hyperactivity and fragmented sleep, older flies experience reduced sleep compared to controls. Conversely, EBV viral particles suppress locomotor activity in young flies while promoting longer and more consolidated sleep.
Reduced activity and increased sleep were detected in EBV DNA or EBV viral particle injected flies during anticipationIn this study, we explored the impact of EBV DNA and EBV viral particles on the anticipatory behaviors of Drosophila melanogaster, specifically during the anticipation phase, which refers to the heightened readiness or activity observed in response to environmental cues such as the scheduled turning on and off of lights. Our primary focus was to assess how these viral injections affected both activity and sleep profiles during the anticipation period, with an emphasis on the underlying regulatory or compensatory mechanisms that may be influenced by EBV exposure.
To evaluate the effects, we analyzed the behavior of the flies during the 6-hour anticipation phase, immediately before the scheduled light changes. Our results indicate that there were no significant differences between the EBV DNA and EBV viral particles groups, nor between EBV DNA and sterile water controls. However, when comparing the EBV viral particle group to its PBS-injected control group during the AM anticipation phase, a significant decrease in activity was observed in the EBV viral particle group (mean = 19.71 min) compared to the PBS group (mean = 66.67 min, p = 0.0012). In the PM phase, we observed lower activity in both the EBV DNA and EBV viral particles injected groups compared to the control groups. Specifically, EBV DNA injected flies exhibited reduced activity (mean = 87.87 min) compared to the sterile water group (mean = 113.75 min), and similarly, EBV viral particle injected flies had lower activity (mean = 84.12 min) compared to their PBS-injected control group (mean = 130.38 min) but not significant (Fig. 4a). Interestingly, the relationship between activity and sleep in Drosophila is not straightforward, as increased sleep does not necessarily correlate with decreased activity. The balance between activity and sleep is influenced by various factors, including circadian rhythms, environmental conditions, and the physiological state of the flies. During both the AM and PM phases, we noted a significantly higher sleep rate in the EBV viral particle group compared to the EBV DNA group (p = 0.0106 and p < 0.0001 respectively). Specifically, in the AM phase, EBV DNA-injected flies had a higher sleep rate (mean = 296.53 min) compared to their control group (mean = 282.97 min, p = 0.025). However, during the PM phase, EBV DNA-injected flies exhibited a significantly lower sleep rate (mean = 190.65 min) compared to the sterile water control group (mean = 296.53 min, p = 0.0001). The EBV viral particle injected flies demonstrated an increased sleep rate during both the AM and PM anticipation periods. In the AM phase, the sleep duration was higher in the EBV viral particles group (mean = 341.23 min) compared to the PBS control group (mean = 282.97 min). In the PM phase, EBV viral particle injected flies also had significantly higher sleep rates (mean = 286.51 min) compared to the PBS group (mean = 239.18 min, p = 0.0048) (Fig. 4b). Finally, when analyzing the number of sleep bouts, the EBV-injected groups exhibited fewer sleep bouts compared to the PBS control group. Specifically, EBV DNA and EBV viral particles injected flies had significantly fewer sleep bouts (mean = 2.38 and 3.86, respectively) compared to the PBS-injected W1118 control flies (mean = 5.48 and 7.71, respectively) during both the AM and PM phases (Fig. 4c). Our results suggest that EBV viral particles have a significant impact on the anticipatory behaviors of fruit flies, influencing both activity and sleep patterns, with notable differences observed between the AM and PM anticipation periods. These findings highlight the complex interactions between viral infection and circadian behavior, warranting further investigation into the underlying regulatory mechanisms at play.
Fig. 4
EBV DNA or Viral Particles Disrupt Anticipation in Drosophila Melanogaster: Reduced Activity and Increased Sleep. (a) Mean average activity of the male flies during the anticipation phase: 6 h before the lights turn on (am) and 6 h before the lights turn off (pm). (b) Average sleep (min) of the flies. (c) Number of sleep bouts was calculated. Kruskal-Wallis was performed; * p < 0.05; ** p < 0.005; *** p < 0.0005; **** p < 0.0001. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 31, control PBS injected W1118:♂n = 32; EBV 70 viral particles injected W1118: ♂n = 22
Arrhythmic behavioral patterns in EBV DNA-injected fliesFruit flies that display regular peaks and troughs over 24 h and consistent, predictable patterns of activity and rest that correspond with their circadian rhythm are said to be rhythmic. Arrhythmic flies, on the other hand, don’t follow this regular pattern; instead, they exhibit erratic or sporadic periods of activity and rest, which suggests that their circadian clock mechanisms aren’t working properly.
Analysis over 30 days revealed that 10% of EBV DNA-injected flies exhibited arrhythmic behavioral patterns characterized by erratic, non-cyclic activity and rest periods (Fig. 5a). Rhythmicity strength, quantified as the power of rhythmicity (Qp), was significantly weaker in EBV DNA-injected flies compared to sterile water-injected controls (mean = 478.926 vs. 887.979, p = 0.0288). In contrast, EBV viral particle-injected flies displayed a significantly stronger rhythmic behavioral pattern than both PBS and EBV DNA-injected flies (mean = 1327.29 vs. 382.443 and 478.926, respectively, p < 0.0001) (Fig. 5b). These findings suggest that EBV DNA disrupts circadian regulation, leading to erratic behavioral patterns, whereas EBV viral particles enhance rhythmicity strength, potentially reinforcing biological clock mechanisms. EBV DNA disrupts circadian rhythmicity in a subset of flies, leading to arrhythmic behavior, whereas EBV viral particles strengthen rhythmic patterns. These findings highlight the differential effects of EBV DNA and viral particles on age-dependent behavioral regulation.
Fig. 5
Rhythmicity patterns in Drosophila melanogaster. (a) Representative graph of percentage of rhythmicity and arrhythmicity of male W1118 injected either with Sterile Water (n = 28), EBV DNA (n = 31), PBS (n = 32) or 70 EBV viral particles (n = 22). (b) Graph illustrating the Qp statistical value (rhythmicity power) of male flies. Kruskal-Wallis was performed * p < 0.05; ** p < 0.005; *** p < 0.0005; **** p < 0.0001. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 31, control PBS injected W1118:♂n = 32; EBV 70 viral particles injected W1118: ♂n = 22)
Activity and sleep disturbances in W 1118 flies injected with EBV DNA or EBV viral particles under LD-DD conditionsGiven the distinct behavioral phenotypes observed in W1118 flies injected with EBV DNA or EBV viral particles, we sought to investigate their role in regulating the endogenous circadian clock by analyzing activity and sleep patterns during the transition from light-dark (LD) to constant dark (DD) conditions (Fig. 6a,c). EBV DNA injected flies exhibited significant activity disruptions under LD conditions, with EBV DNA-injected flies displaying a lower mean activity compared to EBV viral particle-injected flies (mean = 294.655 min vs. 465.407 min, p = 0.028) (Fig. 6b). Upon shifting from LD to DD, EBV DNA-injected flies showed significantly reduced anticipation peaks before ZT0 (morning transition) and ZT12 (evening transition) compared to sterile water-injected controls, with a significant decrease in mean activity during LD (p = 0.0080). In contrast, EBV viral particle-injected flies lacked distinct anticipation peaks but did not show significant variations in activity levels compared to PBS-injected controls during the LD-DD transition (Fig. 6b,d). Sleep profiles differed significantly between EBV DNA and EBV viral particle-injected flies (Fig. 6e,h). EBV DNA-injected flies exhibited significantly higher mean sleep duration compared to both EBV viral particles (p = 0.0103 in LD and p = 0.0454 in DD) and sterile water-injected controls (mean = 1274.74 min vs. 982.536 min, p < 0.0001 in LD; p < 0.0001 in DD). EBV viral particle-injected flies exhibited a distinct sleep pattern, showing a significant reduction in sleep duration compared to PBS-injected controls (p = 0.0328) during LD conditions only (Fig. 6f,i). Sleep fragmentation, measured by sleep bout count, also varied significantly. During LD, a clear distinction was observed between EBV DNA and EBV viral particle-injected flies (p < 0.0001) (Fig. 6g). EBV DNA-injected flies exhibited fewer sleep bouts compared to sterile water-injected controls (mean = 2.85172 vs. 28.2857, p < 0.0001 in LD; mean = 19.931 vs. 32.9643, p = 0.0005 in DD). In contrast, EBV viral particle-injected flies exhibited an increased number of sleep bouts compared to PBS-injected controls during LD (mean = 23.5379 vs. 13.0174, p = 0.0491) (Fig. 6j). These results suggest that EBV DNA disrupts sleep stability while EBV viral particles increase sleep fragmentation under LD conditions.
Fig. 6
EBV DNA and EBV viral particles W1118 injected flies exhibit circadian activity and sleep disruptions. (a-c) Graph showing the average activity profile of all injected groups, with activity counts measured every 5 min over 24 h. By an average of 5 days under light/dark conditions (LD1-LD5) and 5 days under dark/dark conditions (DD1-DD5), respectively. Two activity bouts are centered: ZT0 (morning peak) and ZT12 (evening peak). (b-d) Mean locomotor activity per day of all injected groups obtained by averaging 5 days in light/dark conditions (LD1-LD5) and 5 days in dark/dark conditions (DD1-DD5).(e-h) Graph displaying the average sleep profile of all injected groups, with activity counts measured every 5 min over a 24-hour period by averaging over 5 days under light/dark conditions (LD1-LD5) and 5 days under dark/dark conditions (DD1-DD5).(f-i) Average sleep (minutes) of the flies over 24 h intervals during LD and DD periods. (g-j) Number of sleep bouts calculated during 24 h of spontaneous sleep during LD and DD periods. Kruskal-Wallis was performed as a statistical significance: * p < 0.05; ** p < 0.005; *** p < 0.0005; **** p < 0.0001. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 29; control PBS injected W1118:♂n = 23; EBV 70 particles injected W1118: ♂n = 29). Error bars represent standard deviation
EBV DNA and EBV viral particles affect circadian function in fliesUnder constant environmental conditions, organisms rely on their endogenous circadian rhythm, which follows a free-running cycle. In Drosophila melanogaster, this cycle is approximately 1440 min, and deviations from this period indicate potential circadian clock dysfunction. During the LD phase, both EBV DNA and EBV viral particle-injected flies exhibited a reduced free-running period compared to controls. A total of 51.19% of EBV DNA-injected flies had a free-running period exceeding 1440 min, compared to 81.48% of sterile water-injected controls. Similarly, 72.41% of EBV viral particle-injected flies had a prolonged free-running period, compared to 87.5% of PBS-injected controls (Fig. 7a). However, in the DD phase, these effects varied between the two groups. Only 35% of EBV DNA-injected flies maintained a free-running period above 1440 min, compared to 59.26% of sterile water-injected controls. In contrast, 52.17% of EBV viral particle-injected flies exhibited a prolonged free-running period, while only 4.35% of PBS-injected controls did (Fig. 7b). These findings indicate that while both EBV DNA and viral particles alter circadian timing, the effects are more pronounced under constant dark conditions. The transition from LD to DD also revealed increased arrhythmicity in EBV-injected flies. During LD, 6.90% of sterile water-injected flies and 3.57% of EBV DNA-injected flies exhibited arrhythmic behavior. However, in DD conditions, the arrhythmicity rate remained unchanged in sterile water-injected flies but increased to 13.04% in EBV DNA-injected flies. Similarly, EBV viral particle-injected flies, which exhibited 0% arrhythmicity in LD, showed an 8% arrhythmicity rate in DD (Fig. 7c). These results confirm that both EBV DNA and EBV viral particles impair circadian stability, with effects becoming more pronounced in constant darkness.
Fig. 7
Both EBV DNA and EBV viral particle injections affect the circadian function of flies. (a) Decreased free-running period during LD period both in EBV DNA and EBV particles injected flies. Percentage of flies with a period greater or equal to/ lower than 1440 min during light-dark. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂= 29; control PBS injected W1118:♂n = 23; EBV 70 particles injected W1118: ♂n = 29. (b) Free-running period variability during DD period in both EBV DNA and EBV viral particle injected flies. Percentage of flies with a period greater or equal to/ lower than 1440 min during dark-dark. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 29; control PBS injected W1118: ♂n = 23; EBV 70 particles injected W1118: ♂n = 29. (c) Percentage of arrhythmic flies under LD/DD conditions. Control Sterile water injected W1118: ♂n = 28; EBV DNA injected W1118: ♂n = 29; control PBS injected W1118: ♂n = 23; EBV 70 particles injected W1118: ♂n = 29
Comments (0)