
Remember me

HASIntra was expressed and purified as a ∼30 kDa non-membrane bond protein (Additional file 1: Fig. S3). The purification of membrane proteins is challenging due to their low solubility in conventional detergents. The efficacy of detergent usually depends on the membrane protein type and the purification conditions. For this reason, in the current study, different nonionic detergents including DDM, Triton X-100, and Tween 20 were applied to compare the extent of protein solubilization. In our previous study, we purified the membrane variants using SDS as a surfactant. However, as our data revealed, the loss of protein, as well as the reduction in activity, was remarkable. Data illustrated that all detergents could solubilize the bacterial membrane. However, DDM led to more solubilization (Additional file 1: Table S3). Nonetheless, more solubilization by a detergent may not necessarily means maintaining protein structure and activity. Therefore, the activity of variants was quantitatively evaluated by measuring the amount of HA in vitro synthesis. The concentration of HA in individual reactions, as well as the specific enzyme activity of variants was calculated. Data revealed that the variants purified by Triton X-100 and Tween 20 have a remarkable reduction in activity when compared to that of the DDM-purified (Additional file 1: Table S3). In DDM-based purification, although all variants kept the biosynthesis capability, however, they showed lower specific activity rather than the wild-type (Table 1). HA titer was statistically different among the variants, so that SeHAS and HASIntra had the highest and lowest HA titer, respectively (Additional file 1: Fig. S4). Determination of protein impurities in HA samples demonstrated that all are in the range of acceptance limit according to European Pharmacopoeia 10. The purification steps did not have deleterious effects on HA Mw when tested by the control.
Table 1 HA titer and specific activity of recombinant variants after purification with DDMEnzyme kineticsThe kinetic behaviors of SeHAS variants are shown in Figs. 1 and 2. As noted in previous studies, SeHAS appeared to be more catalytically active than the truncated forms (Cohan et al. 2023; Tlapak-Simmons et al. 2004). The substrate concentration significantly alters the activity of variants (p-value < 0.0001). The activity of SeHAS and truncated forms at different UDP-GlcUA concentrations were 8.5, 5.6, 4.7, and 1.6 µgHA µgenzyme−1 min−1 for SeHAS, HAS123, HAS23, and HASIntra, respectively. Similarly, the activity of SeHAS and truncated forms at different UDP-GlcNAc concentrations were 8.2, 5.5, 3.4, and 1.4 µgHA µgenzyme−1 min−1 for SeHAS, HAS123, HAS23, and HASIntra, respectively. The Km values for both substrates differed slightly at the saturated concentration (Table 2). Statistical analysis indicated significant differences in the Michaelis constants of KmUDP-GlcUA (p-value < 0.0001), suggesting that the variance in the binding affinity of the two UDP-sugars controls the molecular weight of hyaluronan. The Vmax profiles for all enzymes were hyperbolic, while the Lineweaver–Burk plots were linear (Additional file 1: Fig. S5).
Fig. 1
Effect of UDP-GlcUA concentration on the activity of SeHAS and truncated forms. The Km values for UDP-GlcUA were determined by varying the concentration of UDP-GlcUA from 0.05 to 1.5 mM while keeping the other at 1.5 mM. As depicted, all saturation profiles are hyperbolic
Fig. 2
Effect of UDP-GlcNAc concentration on the activity of SeHAS and truncated forms. The Km values for UDP-GlcNAc were determined by varying the concentration of UDP-GlcNAc from 0.05 to 1.5 mM while keeping the other at 1.5 mM. As depicted, all saturation profiles are hyperbolic
Table 2 The Michaelis–Menten constants obtained for SeHAS and the truncated formsFTIR spectroscopyThe structural identity of synthesized polymers was determined by FTIR and compared to that of the control. The spectra analysis elucidated that there is no obvious difference between the spectra of HA control and purified HAs (Fig. 3). A strong absorption peak was observed at 3302 cm−1, which indicates OH and NH bonds. The absorption peak at 2893 cm−1 was related to CH symmetrical and CH2 asymmetrical stretching. The peaks at positions 1617 cm−1, 1562 cm−1, and 1324 cm−1 can be for amides I, II, and III. The absorption peaks at 1081 cm−1 and 1133 cm−1 are typical for carbohydrates and the peak at 1410 cm−1 is assigned to symmetric C–O stretching vibrations (Chen et al. 2019; Gilli et al. 1994).
Fig. 3
FTIR spectroscopy of purified HAs from SeHAS, HAS123, HAS23, and HASIntra along with control
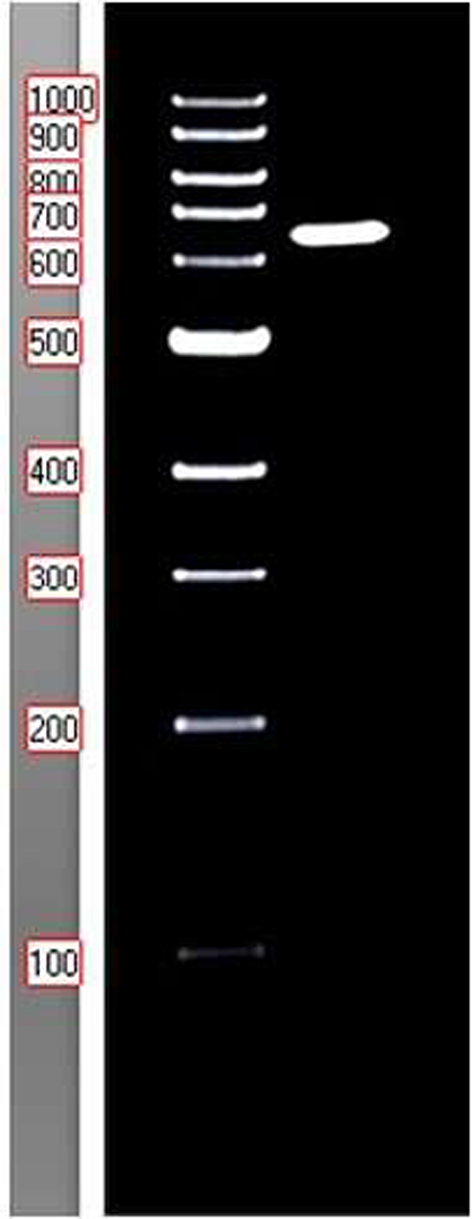
MW and dispersity of polymersThe MW of HAs was determined by PAGE and Alcian blue/silver staining. SeHAS produced a HMW polymer (> 270 kDa), meanwhile HAS123, HAS23, and HASIntra produced LMW ones (< 30 KDa) (Fig. 4). The dispersity of HA polymers is summarized in Table 3. DLS experiments showed the presence of a low-disperse polymer (PDI < 0.3) for HASIntra product (HAIntra) and a polydisperse HA (PDI range: 0.5–1.0) for other products.
Fig. 4
MW determination of HA products [Lane 1: DNA ladder 1 kb, Lane 2: DNA ladder 100 bp, Lane 3: HA ladder (a range from 509 to 30.6 KDa), Lane 4: HA produced by HASIntra, Lane 5: HA produced by HAS23, Lane 6: HA produced by HAS123, and lane 7: HA produced by SeHAS]
Table 3 Dispersity and MW of produced HAs in this studyEndothelial cell proliferation assayHUVECs were stimulated by various concentrations of the synthesized polymers. EC proliferation was negligible after 48 h for all LMW-HAs (data not shown). While, a reverse dose-dependent cell proliferation was occurred after 72 h by LMW-HAs (Fig. 5A). The highest proliferation was for HAIntra and HA10kDa (LMW-HA control) at concentration of 6 µg/mL. In contrast, HASeHAS and HA760kDa (HMW-HA control) showed an inhibitory effect at 150 µg/mL, while lower concentrations (100, 70, and 20 µg/mL) did not affect the EC proliferation (Fig. 5B).
Fig. 5
Cell proliferation assay of synthesized HAs. ECs were exposed to LMW- and HMW-HAs at concentrations of 6–40 µg/mL (A) and 20–150 µg/mL (B), respectively. Data were expressed as mean ± SD from three independent experiments. Statistical comparisons were performed between the lowest concentration relative to the highest concentration (*p-value < 0.05; **p-value < 0.01)
In vitro cell migration assayThe scratched cells were treated with HA samples at two doses (10 μg/mL and 200 μg/mL) and the lesion closing rates were monitored for 24 h. The results showed that the scratched area began to close at 24 h after the addition of exogenous 10 μg/mL LMW-HAs/ HA10kDa, which was in parallel with the EC proliferation test (Fig. 6A). Whereas in other groups (HA760kDa and culture medium), the lesion did not fully close after 24 h (Fig. 6B). Comparing the two doses showed that the lesion healed faster at the lower dose in both LMW-HAs and HMW-HAs groups. The cell imaging demonstrates that the wound undergoes negligible proliferation after treatment with a high concentration of HAs (independent of size) (Additional file 1: Fig. S6).
Fig. 6
Cell migration assay. ECs were exposed to the produced HAs at concentrations of A 10 µg/mL and B 200 µg/mL. The graph reports the percentage of wound closure relative to time zero. Data were expressed as mean ± SD from three independent experiments
DiscussionThe MW of HA dictates its biological function (Ebrahimi et al. 2022; Garantziotis and Savani 2019). Therefore, there is great interest in understanding the mechanisms and factors influencing the size of HA. Moreover, the cell signaling capabilities of LMW-HA have encouraged producers to synthesize the lower mass of the polymer (Simpson et al. 2015). In addition to the generation of smaller HA by degradation of the long-chain polymers, HAS itself could be regulated directly to synthesize smaller HA (e.g. 40–100 kDa) (Baggenstoss et al. 2017; Hascall et al. 2014; Jokela et al. 2011; Moretto et al. 2015; Yang et al. 2017). Despite basic knowledge about HAS, however, major questions regarding the mechanism of synthesis and control of size have still remained unanswered. In total, HAS has intrinsic properties that influence both the rate and size of HA. Thus, these two functions could be regulated and controlled by separate sub-mechanisms (Baggenstoss et al. 2017). Sequence alterations as an intrinsic factor play a role in Mw variations so, the enzyme could be regulated to directly synthesize LMW-HA (Schulte et al. 2019). The hypothesis for the present study is that TMD deletions probably by altering the enzyme structure could control the HA size.
In the current study, we truncated the SeHAS enzyme to make a more detailed investigation of the function of the transmembrane domains. Extended deletion methodology was used to investigate the effects of TMDs on the preparation of LMW-HA at in vitro conditions.
The pioneering work of Dorfman and co-workers in the 1950s and 1960s showed that the streptococcal HAS is located in the cell membrane, requires Mg2+ ions, and uses two sugar nucleotides (UDP-GlcUA and UDP-GlcNAc) to polymerize a HA chain (Markovitz et al. 1959; Stoolmiller and Dorfman 1969). All of the known enzymes catalyze reactions that use one or two (or, rarely, three) substrates and produce one or two products. However, hyaluronan synthases (HASs) are different from the characterized enzymes. HASs possess dual glycosyltransferase activities within a single protein, (β1, 4-GlcNAc transferase and β1, 3-GlcUA transferase), and the hyaluronic acid (HA) product sequentially serves as the acceptor for each subsequent sugar addition. Although it seems straightforward, the enzyme must possess at least six (and probably seven) different functions to perform the overall reaction for the synthesis of one disaccharide unit. HAS enzymes possess an additional distinctive feature, as the structural similarity between the two sugar nucleotides affords each the potential to engage in competitive binding for the appropriate UDP-sugar-binding site on the enzyme. Although the HASs do not misincorporate other sugar nucleotides into the growing HA chain, UDP-GlcUA can interact with the UDP-GlcNAc binding site (Tlapak-Simmons et al. 1999b). Initial experiments have indicated that with the two normal substrates, a significant disparity in the concentration of one UDP-sugar compared to the other leads to variations in the rate of hyaluronic acid (HA) synthesis (Tlapak-Simmons et al. 1999b). There are no previous reports of this cross-talk phenomenon affecting HA biosynthesis between SeHAS and their truncation variants. The observation of this kinetic behavior reflects the advantage of studying purified SeHAS truncation forms and catalytic efficiency comparison between enzyme forms. Here, for the first time, we have characterized the kinetic behavior of purified SeHAS truncated forms. The catalytic region for SeHAS was reported in 264 AA from positions 55 to 319. In the present study, the SeHAS variants were truncated from the N-terminal and C-terminal extremity based on the folding structure. Studies have shown that the intracellular domain has an important role in polysaccharide synthesis. Polysaccharide-synthesizing capability of this domain could be retained depending on the truncation size. The result showed the enzymes displayed a decreasing activity depending on the size of truncation. Our data were similar to the previous studies in that the manipulated enzymes could still produce HA polymer, but differ in the amount of synthesized HA due to differences in the enzyme activity.
Indeed, HAS enzymes are highly lipid-dependent and are most effectively stimulated by cardiolipin. Tlapak-Simmons conducted a study on the influence of cardiolipin on the kinetics of KmUDP-GlcNAc in the purified SeHAS. In the absence of cardiolipin, the purified SeHAS was active and had responses to increasing substrate concentration (Tlapak-Simmons et al. 1999a). A significant enhancement in the synthase activity observed in the presence of cardiolipin may be attributed to a cardiolipin-dependent influence on the enzyme, resulting in alterations in the Km values (four–ten-fold increase) for the sugar nucleotide substrates. They also reported the KmUDPGlcUA and KmUDP-GlcNAc values respectively as 274 µM and 251 µM for detergent-solubilized SeHAS in the presence of 2 mM bovine cardiolipin. They found a hyperbolic behavior for purified SeHAS in response to increasing substrate concentration as shown in our study. However, the reported Km values are much higher than our obtained values. Therefore, the addition of bovine cardiolipin can be explained by such an increase in the specific activity of purified SeHAS. This conclusion is supported by the results of this study showing that cardiolipin-depleted SeHAS, still has enzyme activity. Furthermore, the results demonstrated that the n-dodecyl-d-maltoside-solubilized purified variants were active. This is an important finding because many studies have reported that HAS activity is irreversibly lost upon solubilization of the protein in a wide variety of nonionic detergents (Tlapak-Simmons et al. 1999b).
Interestingly, two substantial differences were observed between SeHAS and the truncated forms. First, the SeHAS is intrinsically more active than the truncated ones. This was apparent in the current and the earlier studies (Cohan et al. 2023; Keramati et al. 2022) that were conducted with both membrane-bond and purified enzymes. With the addition of UDP-sugars, truncated enzymes underwent a glycosylation reaction and transferred the substrate to the acceptor effectively. Second, the truncated variants displayed increased Km values for two substrates when compared to the wild-type enzyme. SeHAS without any deletion showed lower Km values, whereas HASIntra with completely deleted transmembrane domains showed a significantly higher Km value. Kinetic analysis showed that both Km and Kcat of HASIntra were affected significantly. This is an effective way to make use of HASIntra as the shortest enzyme form for HA biosynthesis. The intracellular region has a key role in maintaining SeHAS activity. These results indicate that the presence of TMDs is essential for substrate-binding affinity and deletion of each domain increases the Km value. The kinetic analysis indicated that the low affinity for substrate in the truncated forms regulates the hyaluronan molecular weight and will have a significant effect on the polydispersity of the products (Kooy et al. 2014). Furthermore, these results offer several possible strategies to decrease the polydispersity of the products including metabolic engineering to change the substrate concentration in HA-producing microorganisms and/or single mutation studies to regulate the binding affinity of the substrate within the HAS enzyme.
Multiple groups have created streptococcal HAS mutants to identify property-influencing sites such as activity and HA size (Baggenstoss et al. 2017). Mutational studies involving point-mutation within TMDs and the C-terminus of SeHAS showed different effects on HA synthesis and/or product size (Kumari et al. 2006, 2002). It was demonstrated disrupting the interaction between TMD2 and TMD4 caused the synthesis of smaller polymer (0.6–3.2 MDa versus 3.6 MDa) (Kumari et al. 2006). Some studies examined site-specific mutations in the C-terminus of SeHAS to evaluate the role of this region in the activity and MW control (Heldermon et al. 2001). Mutation’s study showed that polymerizing activity can decouple from the size control. In this regard, a tandem motif region (K398–X7–R406–X7–K414 motif) at the C-terminus of the enzyme was deciphered. They find almost R406 mutants synthesize a larger polymer and the variants show specific activities from 70 to 177% of the native enzyme. While the K398 mutants were associated with a remarkable reduction in the specific activity (14–64% of wild type) that led to a smaller size of polymer (≤ 250–480 kDa) (Baggenstoss et al. 2017). In a similar study, mutations in the last nine residues at the C-terminal of SeHAS (409–417) showed a decrease in the HA titer (16.8% of WT) and MW (< 790 kDa), emphasizing the importance of the C-terminus region for HAS activity and size regulation (Yang et al. 2017). Moreover, this study demonstrates how R406-R413 residues constituted an HA-binding pattern that stabilizes the HA-SeHAS complex and helps the enzyme to regulate the production rate and molecular size of the polymer. They increased the HA product size via site-directed mutation at the C-terminal (residues 414 to 417) based on the hypothesis that higher binding affinity between the SeHAS C-terminus and HA would lead to a larger HA size. A three-fold increase in the size (HA MW = 1270 kDa) due to enhanced binding affinity in the K414R variant suggested that residues 414–417 are involved in polymer retention to make a longer chain (Yang et al. 2017). In our previous study, we focused on the role of two C-terminus TMDs (TMD4 and TMD5) of SeHAS in the recombinant Bacillus subtilis strain and find out that the deletion of these transmembranes did not affect the MW of polymer (Amjad Zanjani et al. 2022).
It was found that C-terminal end deletion has no effect on HA synthesis activity, but it will improve the stability of the enzyme-HA protein complex and lead to the production of HMW-HAs. However, these studies have been limited to the C-terminus region of SeHAS. Therefore, in the current study, we employed extended TMD deletions to clarify the role of other domains in the activity of enzyme and the HA size. Data analysis revealed that TMD deletions alter the activity and polymer size. Following the deletion of all TMDs, the polysaccharide synthesis capability of the shortest form of the enzyme (HASIntra) was significantly preserved. The highest level of activity among the variants was for HAS123 (91% of WT). The results indicated that TMD4, TMD5, and the extracellular C-terminus region have no deleterious effects on HA synthesis. The results confirm earlier findings by Baggenstoss et al. and Yang et al. that C-terminus mutants are able to synthesize HA (Baggenstoss et al. 2017; Yang et al. 2017). SeHAS truncation by Baggenstoss et al. revealed that the deletion of residues 398–417 caused an undetectable HA MW. It is presumed that this mutant is unstable (not expressed), indicating that the HAS C-terminal region is likely critical for enzyme stability (Baggenstoss et al. 2017). In previous studies, the transmembrane topology of SeHAS revealed that the C-terminal peptide of SeHAS from residues 402 to 417 is located inside the cell (Yang et al. 2017). In this study, the topological prediction showed that the C-terminus of GGS-HAS is an extracellular domain, in which deletion of this region may have a different role in the enzyme activity and control of MW of polymer compared to similar studies.
Deletion of first TMD in HAS23 reduces the enzyme activity to 66% of WT. The comparison of HAS123 and HAS23 productivity demonstrated that, in contrast to HAS123, deletion of TMD1 has a significant effect on the HAS activity. It should be notified that in our previous work, we prepared.
HASIntra, HAS123, HAS23, HAS3, and HAS2 variants and preliminary screening of activity elucidated the importance role TMD1, TMD2, and TMD3 for full activity (Cohan et al. 2023). One significant difference in the current work is the use of different purification methods for the variants that leads to obtaining higher specific activity for the variants. We used a hybrid purification method for HASIntra, in which the resin is washed by graduate reduction of urea. This procedure resulted in better folding of the variant and its increased activity (38% versus 11%). For membrane variants, we tested different nonionic detergents such as DDM, Triton X-100, and Tween 20 to evaluate their effects on protein solubilization and activity. As our data demonstrated, among detergents, DDM showed the best results (the highest solubilization and activity) for the variants. It must be notified that the type of detergent is very important for isolation of membrane proteins and should be tested for each protein (Orwick-Rydmark et al. 2016). For example, in a study conducted by Lee et al., different detergents including DDM, Brij-35, Triton X-100, cholate, CHAPSO, Zwittergent 3–12, Deoxy BIG CHAP, and digitonin were used for purification of cadherin-11 and it was found that Triton X-100 and DDM were more efficient than the others (Lee et al. 2018). Similar to our previous work, HASIntra (the central domain of SeHAS) showed a reduction in activity by up to 38%. Although this region as a single functional glycosyltransferase domain is able to bind to the substrates and catalyze the polymerization of HA but requires TMD1, TMD2, and TMD3 for full activity. In general, the activity measurements indicated that these TMDs are essential for accelerating the synthesis of the polymer.
Despite our previous study that clarified the effect of TMDs on the SeHAS activity, however, the role of the N-terminal region on the control of product size was not investigated so far. Therefore, in the current study, the effect of such deletions on the polymer size was assessed. Our results illuminated that in comparison to SeHAS, which produced a polydisperse HMW polymer, HAS123 and HAS23 produced polydisperse LMW-HAs. Meanwhile, HASIntra produced a low-disperse LMW-HA. Preparation of smaller HA products (< 30 kDa) by the variants compared to SeHAS may be due to the removal of TMDs in the N- and C-terminal of the enzyme that probably alters the conformation of the enzyme. The HA Mw produced by HAS123 and HAS23 was almost identical with very similar polydispersity. This result is not unexpected, since HAS123 and HAS23 show variation at only a position, including TMD1. Maybe it can be concluded that the first TMD may be more involved in the synthesis rate than the size control.
Of the biological activities of HA, the most interesting one is the angiogenesis capability of LMW-HA in wound repair (cell migration assay) through stimulating EC proliferation (Mo et al. 2011). Therefore, to test the biological activity of produced polymers, a cell proliferation assay was utilized. We observed all produced LMW-HAs (HA123, HA23, and HAIntra), as well as the control (HA10kDa) could stimulate EC proliferation in a dose-dependent manner with maximum value at 6 µg/mL after 72 h. The data also elucidated that this stimulatory effect was highest for HAIntra and HA10kDa, which are low-disperse. This phenomenon highlights the importance of LMW low-disperse preparations of this polymer for medicinal applications. On the contrary, HASeHAS and HA760kDa (control) showed only an inhibitory effect at 150 µg/mL without any stimulatory effect at lower concentrations. Previous studies evaluated the regulatory effect of HAs on HUVEC proliferation and indicated that ECs respond poorly to large HAs with a reduction in the growth at 100–500 µg/ml. While LMW-HAs have stimulatory effects on these cells at 3–20 μg/mL by interacting with receptor for HA-mediated motility (RHAMM) and CD44 as receptors (Ibrahim and Ramamurthi 2008; Mo et al. 2011; Queisser et al. 2021). In addition to the proliferation assay, we investigated the biological activity of polymers by a cell migration assay in vitro model (Gao et al. 2008). Interestingly, LMW-HAs were fully active for cell proliferation and migration. The cell migration test demonstrated that HA123, HA23, HAIntra, and HA10kDa fully recover the lesion after 24 h at the lower dose (10 µg/mL). Nonetheless, in the case of HASeHAS and HA760kDa, the scratched area did not fully close. Cell imaging also demonstrated that an increase in the HA concentration postponed the healing of HUVEC, as all groups, independent in size, did not show a full recovery within 24 h. These results are in agreement with the literature data that LMW-HAs are strong inducers of angiogenesis and recover the EC wound with different origins at low concentrations (Gao et al. 2006, 2008; Mo et al. 2011; Slevin et al. 2002; Wang et al. 2016). Our data also confirms the opposing effects of LMW-HA at low (stimulatory) and high concentrations (inhibitory) in wound recovery (Gao et al. 2
Comments (0)