
Remember me
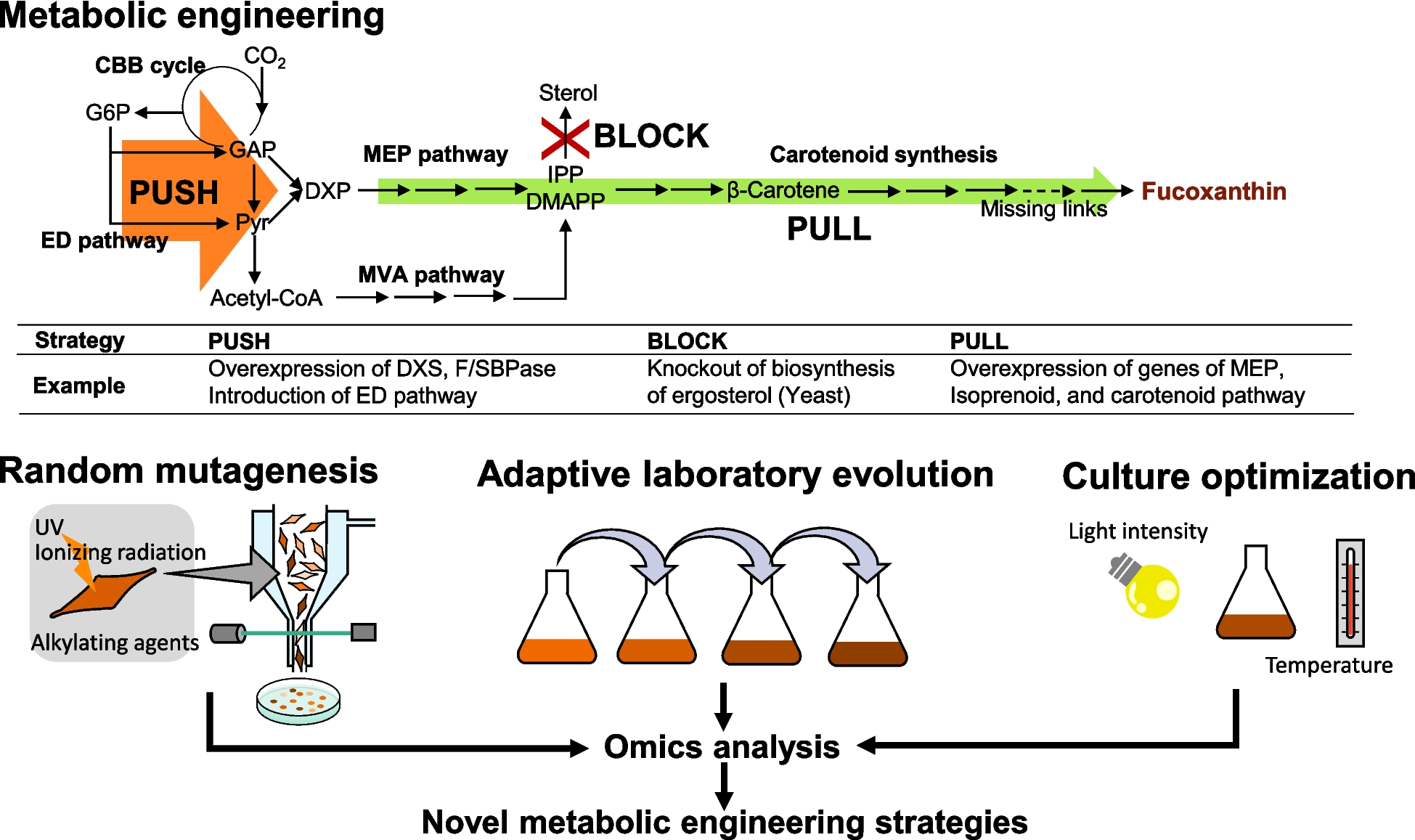



Firstly, the effect of cysteine on the growth of the C. glutamicum wild-type NBRC 12168 in the modified M9 medium was analyzed (Fig. 1a). In this study, the medium that does not contain natural components such as peptone and yeast extracts was used to evaluate the effect of cysteine on cell growth because the effect of unknown nutrients in natural components needs to be avoided. In the wild-type strain, cysteine inhibited its growth and the degree of growth inhibition was dose-dependent when adding 0.25–1 g L−1 Cys·HCl·H2O to the medium; the specific growth rate during exponential growth decreased as the cysteine concentration added to the medium increased (Supplementary Table S1). However, cell growth was not completely inhibited by the addition of 0.5–2 g L−1 Cys·HCl·H2O.
Fig. 1
Effect of cysteine on the growth of C. glutamicum. Cell growth of the C. glutamicum NBRC 12168 (a) and its aecD gene disruptant (b) in modified M9 medium without cysteine addition (black circles) and with the addition of 0.25 (red triangles), 0.5 (blue squares), and 1 (green diamonds) g L−1 of Cys·HCl·H2O is shown. The average ± standard deviation of the data in three independent cultures is shown
Next, the effect of cysteine on the growth of the disruptant of the aecD gene encoding cysteine desulfhydrase, which is responsible for cysteine degradation, was analyzed (Fig. 1b). The growth of the aecD disruptant was dramatically reduced even in the presence of 0.25 g L−1 Cys·HCl·H2O (Fig. 1b). Similar to the result in the NBRC 12168, the specific growth rates reduced as the cysteine concentration added to the medium increased (Supplementary Table S1). The results indicate that the aecD disruptant exhibits more sensitivity to cysteine than the wild-type strain because the aecD disruptant cannot degrade cysteine incorporated into cells.
ALE of C. glutamicum in the presence of cysteineIn order to obtain cysteine-resistant mutants of C. glutamicum, ALE in the presence of cysteine was performed. In the present study, the aecD gene disruptant was used to avoid degrading cysteine by the cells during ALE. Here, five independent ALE series, namely ALE1, ALE2, ALE3, ALE4, and ALE5, were conducted. The growth profiles of five ALE experiments are shown in Supplementary Fig. S1. Cell growth in all five ALE series was finally observed in a modified M9 medium containing 4 g L−1 Cys·HCl·H2O after 2200–2300 h, which corresponds to 330–370 generations, and the specific growth rate reached approximately 0.13 h−1.
To confirm whether the cultures obtained at the end of the ALE actually contain cells that can grow in the presence of cysteine, growth of the cell populations at the end of the ALE in the medium containing 4 g L−1 Cys·HCl·H2O was measured. As shown in Supplementary Fig. S2, cell populations in all ALE cultures could grow well in the presence of 4 g L−1 Cys·HCl·H2O. This result indicates that cell populations in the ALE cultures contain mutants that can grow in the presence of 4 g L−1 Cys·HCl·H2O.
Furthermore, to confirm whether the cell populations in the ALE cultures stably exhibited cysteine resistance, serial transfer of the ALE cultures to the modified M9 medium without cysteine addition was repeated five times. The cultures were then transferred to the same medium containing 4 g L−1 Cys·HCl·H2O, and the growth was measured. As shown in Supplementary Fig. S3, cell populations of two ALE culture lineages, ALE2 and ALE5, showed higher growth in the presence of 4 g L−1 Cys·HCl·H2O compared to those of the other ALE culture linages, ALE1, ALE3, and ALE4. Therefore, ALE2 and ALE5 were chosen to isolate cysteine-resistant mutants of C. glutamicum because it was expected that the genomes of the cells in these ALE cultures have mutations related to cysteine resistance.
Isolation of cysteine-resistant mutants of C. glutamicum from ALE culturesThe cysteine-resistant mutants of C. glutamicum were obtained from the cultures of ALE2 and ALE5 obtained at the end of ALE experiments (2279 h; 362 and 352 generations, respectively) as described in the “Materials and methods” section. Among the ten strains, four and five strains from ALE2 and ALE5, respectively, showing the highest specific growth rates in the presence of 4 g L−1 Cys·HCl·H2O, namely ALE2_1 (0.12 h−1), ALE2_2 (0.13 h−1), ALE2_4 (0.14 h−1), and ALE2_10 (0.12 h−1) from ALE2 and ALE5_2 (0.14 h−1), ALE5_6 (0.12 h−1), ALE5_7 (0.15 h−1), ALE5_9 (0.12 h−1), and ALE5_10 (0.12 h−1) from ALE5, were selected and cultured again to measure their specific growth rates (Supplementary Fig. S4). As a result, two strains showing the highest specific growth rates in the presence of 4 g L−1 Cys·HCl·H2O from each of ALE2 and ALE5, namely ALE2_1 (0.12 h−1) and ALE2_10 (0.15 h−1) from ALE2 and ALE5_2 (0.16 h−1) and ALE5_10 (0.11 h−1) from ALE5, were selected.
Further, ten colonies from the four selected strains grown on L agar plates were cultured again in the modified M9 medium containing 4 g L−1 Cys·HCl·H2O to measure their growth. In the case of the strains obtained from ALE2_10, four strains, ALE2_10_3, ALE2_10_6, ALE2_10_7, and ALE2_10_8, exhibited the highest specific growth rate in the presence of 4 g L−1 Cys·HCl·H2O; their specific growth rates were 0.13, 0.13, 0.13, and 0.14 h−1, respectively. Among them, the ALE2_10_3 strain showed the highest OD660 value at 12 h, which corresponds to the initial increase of cell growth after starting the cultivation, and was selected as a cysteine-resistant strain for further analysis (Supplementary Fig. S4a). Moreover, ALE5_2_9 and ALE5_10_4 were selected from the ALE5_2 and ALE5_10, respectively, as cysteine-resistant mutants for further analysis (Supplementary Fig. S4b), as they demonstrated specific growth rates of 0.15 and 0.16 h−1, respectively. No strains were selected for further analysis from ALE2_1, as no strains exhibited a higher specific growth rate higher than that of the original ALE2_1 (approximately 0.12 h−1) in the presence of 4 g L−1 Cys·HCl·H2O (Supplementary Fig. S4a).
For confirmation of cysteine resistance of the isolated mutants, ALE2_10_3, ALE5_2_9, and ALE5_10_4 strains cultured in the presence of 4 g L−1 Cys·HCl·H2O were inoculated into the same medium and cultured for 24 h. As shown in Fig. 2, all the mutants grew well in the presence of 4 g L−1 Cys·HCl·H2O, and specific growth rates of the ALE2_10_3, ALE5_2_9, and ALE5_10_4 strains during exponential growth (12–24 h) were 0.14 ± 0.01, 0.13 ± 0.001, and 0.13 ± 0.002 h−1, respectively. From the results, the cysteine resistance of three strains was successfully confirmed.
Fig. 2
Cysteine resistance of the mutants obtained from the aecD gene disruptant of C. glutamicum via ALE. Cells cultured in the absence (the aecD gene disruptant) or presence (the isolated mutants) of 4 g L−1 Cys·HCl·H2O for 24 h were grown in a modified M9 medium containing 4 g L−1 Cys·HCl·H2O to confirm cysteine resistance of the isolated mutants. Cell growth of the aecD disruptant (black circles) and the mutants ALE2_10_3 (red triangles), ALE5_2_9 (blue squares), and ALE5_10_4 (green diamonds) is shown. The average ± standard deviation of the data in three independent cultures is shown
Identification of mutations related to cysteine resistance in C. glutamicumTo identify the mutations responsible for cysteine resistance in the cysteine-resistant mutants obtained by ALE, whole genome resequencing of the cell populations in ALE cultures and cells of the cysteine-resistant mutants was conducted. Here, the genomic DNA obtained from the cell populations from ALE2 and ALE5 at the end of ALE experiments (2279 h; 362 and 352 generations, respectively), the cysteine-resistant mutants of ALE2_10_3, ALE5_2_9, and ALE5_10_4, and the aecD disruptant was resequenced. In addition, the genome of the cell populations from ALE2 at 1655.8 h (248 generations) and ALE at 1680.5 h (240 generations) was also resequenced to compare the mutations in the cell populations just before increasing specific growth rates with those at the end of the ALE. The summary of the genome resequencing analyses is provided in Supplementary Table S2.
Genome resequencing analysis revealed mutations in the ilvN (NCgl1223) gene were detected in the genomes of both ALE2 and ALE5 cell populations and all the cysteine-resistant mutants isolated from the ALE cultures (Table 1). The ilvN gene encodes the regulatory small subunit of AHAS, which is involved in the biosynthesis of branched-chain amino acids (Keilhauer et al. 1993). The mutation found in the ALE2_10_3 strain, replacement of the 41st serine residue with proline (S41P) in the ilvN gene product, was also detected in cell populations from ALE2 at 248 and 362 generations. In contrast, the mutation found in the ALE5_2_9 and ALE5_10_4 strains, replacement of the 15th valine residue with isoleucine (V15I) in the ilvN gene product, was not detected in the cell population from ALE5 at 240 generations but was detected at 352 generations. The mutation, replacement of the 17th aspartic acid residue with glutamic acid (D17E) in the ilvN gene product, was found in both 240 and 352 generations of the ALE5 lineage. These results suggest that the mutations in the ilvN gene corresponding to S41P and D17E substitutions seem responsible for adaptation to the conditions where the cells grow in the presence of 1.5–3 g L−1 Cys·HCl·H2O.
Table 1 Mutations found in the ilvN gene of the ALE cell populations and isolated cysteine-resistant mutantsTo clarify which mutation(s) in the ilvN gene were related to cysteine resistance in the cysteine-resistant mutants, each identified mutation was introduced into the ilvN locus in the genome of the aecD gene disruptant and the growth of the constructed strains in the presence of cysteine was evaluated. As shown in Fig. 3a–d, the introduction of the ilvN mutations corresponding to S41P and D17E resulted in improved cell growth in the presence of 0.5 g L−1 Cys·HCl·H2O, while the introduction of the other ilvN mutation corresponding to V15I improved the growth slightly. These results indicate that the ilvN mutations S41P and D17E are responsible for resistance to cysteine in C. glutamicum.
Fig. 3
Effect of introduction of ilvN mutations into the aecD gene disruptant of C. glutamicum on growth and valine production in the presence of cysteine. Cell growth of the aecD disruptant (a) and the aecD disruptant carrying the mutations S41P (b), D17E (c), and V15I (d) in ilvN gene in modified M9 medium without cysteine addition (black circles) and with the addition of 0.5 g L−1 Cys·HCl·H2O (red triangles) is shown. In addition, valine concentration in culture supernatant obtained from the culture experiments at 24 h in (a)–(d) in the absence (black bars) and presence (red bars) is shown (e). The average ± standard deviation of the data in three independent cultures is shown
Effect of branched-chain amino acids on growth in the presence of cysteine in C. glutamicumThe concentration of branched-chain amino acids in the culture supernatant obtained at 24 h, shown in Fig. 3a–d, was quantified because the ilvN gene product is involved in branched-chain amino acid biosynthesis. As shown in Fig. 3e, valine was detected in the culture supernatants of strains carrying the ilvN mutations S41P and D17E both with and without the addition of cysteine, whereas for the strain carrying the ilvN mutation V15I, valine was detected in the supernatant without cysteine addition, but this was not observed in the presence of cysteine. The other branched-chain amino acids, leucine and isoleucine, were not detected in the culture supernatants of any of the strains examined (data not shown). These results indicate that ilvN mutations are related to increased valine formation in C. glutamicum.
It can be speculated that the enhancement of valine formation by ilvN mutations mitigates growth inhibition by cysteine in C. glutamicum. Therefore, the effect of valine on the growth inhibitory effect of cysteine in C. glutamicum was investigated. As shown in Fig. 4, the addition of 0.1 g L−1 valine improved the cell growth of the aecD gene disruptant in the presence of 0.5 g L−1 Cys·HCl·H2O, and further improvement of growth was observed upon 0.2 g L−1 valine addition. The addition of leucine slightly improved the growth in the presence of Cys·HCl·H2O, while the addition of isoleucine did not. These results indicate that valine can mitigate the growth inhibitory effect of cysteine in C. glutamicum, and that leucine can slightly suppress it.
Fig. 4
Effect of branched-chain amino acids on growth of the aecD gene disruptant of C. glutamicum in the presence of cysteine. Growth of the aecD gene disruptant of C. glutamicum in modified M9 medium without cysteine addition (black circles) and with addition of 0.5 g L−1 Cys·HCl·H2O alone (red triangles) and 0.5 g L−1 Cys·HCl·H2O plus 0.1 g L−1 (blue squares) or 0.2 g L−1 (green diamonds) valine (a), leucine (b), or isoleucine (c) is shown. The average ± standard deviation of the data in three independent cultures is shown
Effect of ilvN mutations on amino acid production in C. glutamicumTo investigate the effect of ilvN mutations on the biosynthesis of amino acids other than branched-chain amino acids, amino acids in the culture supernatant of the ilvN mutation-carrying aecD gene disruptant were measured (Supplementary Fig. S5). Since the detection sensitivity of the measurement system for branched-chain amino acids described above was low, we used another precolumn amino acid measurement system with high detection sensitivity.
Using this measurement system, leucine could be detected in the culture supernatant of the aecD gene disruptant carrying the mutant ilvN genes, whereas it was not detected in the culture supernatant of the parental aecD gene disruptant. This result indicates that the ilvN mutations result in increased production of leucine as well as valine. However, its production levels were lower than those of valine production. Moreover, glycine production was detected as well in the ilvN mutation-carrying aecD gene disruptant whereas it was not detected in the aecD gene disruptant, suggesting that the ilvN mutations might affect glycine biosynthesis. In addition, histidine production by the aecD gene disruptant was detected, but it was not detected in the ilvN mutation-carrying aecD gene disruptant, suggesting that ilvN mutations might negatively affect histidine biosynthesis.
The katA gene encoding catalase is not related to the mitigation of growth inhibitory effect by valineAs described in the “Introduction” section, cysteine is related to the induction of oxidative stress by the Fenton reaction because it can reduce Fe3+ to Fe2+. In the Fenton reaction, hydroxyl radical is produced from hydrogen peroxide using Fe2+ as an oxidant. If valine induces the synthesis of catalase, which catalyzes the degradation of hydrogen peroxide to water and oxygen, and synthesized catalase enhances the degradation of hydrogen peroxide, the chance to occur the Fenton reaction with Fe2+ produced by the reduction of Fe3+ with cysteine would be reduced, and as a result, growth inhibitory effect of cysteine would be mitigated. Therefore, we investigated the effects of valine addition and ilvN mutations on katA gene expression in C. glutamicum with RT-qPCR.
As shown in Supplementary Fig. S6, the katA gene expression was not significantly changed by the addition of 0.1 g L−1 valine in the aecD disruptant of C. glutamicum. Moreover, the katA gene expression in the aecD disruptant carrying ilvN S41P mutation was lower than that in the parental aecD disruptant, whereas the katA gene expression in the aecD disruptant carrying ilvN D17E and V15I mutations was not significantly different from that in the parental aecD disruptant (Supplementary Fig. S6). These results indicate that catalase does not seem related to the mitigation of growth inhibition by cysteine in C. glutamicum.
Moreover, the effect of katA gene disruption on the mitigation of the growth inhibitory effect of cysteine by valine was investigated. As shown in Supplementary Fig. S7, valine could improve the decreased growth in the presence of 0.5 g L−1 Cys·HCl·H2O even though the katA gene was disrupted in the aecD disruptant of C. glutamicum. This result indicates that the mitigation of the growth inhibitory effect of cysteine by valine is not related to oxidative stress in C. glutamicum.
Effect of cysteine on activity of AHAS from C. glutamicumIt could be speculated that growth inhibition cysteine was caused by inhibition of AHAS activity which resulted in perturbation to valine biosynthesis. Therefore, the activity of AHAS from the aecD gene disruptant in the presence of cysteine was measured. As shown in Supplementary Fig. S8, the activity of AHAS from the aecD disruptant was reduced by about 40% with 0.1 g L−1 Cys·HCl·H2O, indicating that cysteine inhibits AHAS activity in C. glutamicum. Considering this result, the growth inhibitory effect of cysteine in C. glutamicum is related to the inhibition of AHAS activity by cysteine. As expected, the AHAS activity was inhibited by 100 mM valine.
Moreover, the activity of AHAS from the aecD disruptant harboring ilvN mutations in the presence of valine and cysteine was also investigated. As shown in Supplementary Fig. S8, the activity of AHAS from the aecD disruptant carrying ilvN mutations (i.e., S41P, D17E, and V15I) was not inhibited by valine, indicating that this phenomenon is consistent with enhanced valine production in the aecD gene disruptant carrying ilvN mutations. Interestingly, the activity of AHAS from the aecD disruptant carrying the ilvN S41P mutation was inhibited by 0.1 g L−1 Cys·HCl·H2O, but more than 80% of the activity remained. Similarly, 0.1 g L−1 Cys·HCl·H2O inhibited the activity of AHAS from the aecD disruptant carrying the ilvN mutations D17E and V15I, but around 70% of the activity remained. These results indicate that AHAS containing the mutant IlvN protein as a small subunit exhibits resistance to cysteine, and this phenomenon is caused by cysteine resistance of the ilvN mutation-carrying aecD gene disruptant of C. glutamicum.
Cysteine production by engineered cysteine-resistant mutantsFinally, the effectiveness of cysteine-resistant mutants obtained by ALE as bioproduction hosts was investigated. In the present study, cysteine production by engineered cysteine-resistant mutants was examined. The plasmid pCYS*_Cg(T94A)-SER* carrying the mutant cysE and serA genes from C. glutamicum (Kondoh and Hirasawa 2019) was introduced into the cysteine-resistant mutants ALE2_10_3, ALE5_2_9, and ALE_5_10_4 as well as the parental aecD gene disruptant, and cysteine production by the constructed recombinant strains was evaluated. In the experiments, yeast extract was added to the production medium because the addition of yeast extract enhances cell growth and cysteine production as reported previously (Kondoh and Hirasawa 2019).
Cell growth of the cysteine-resistant mutants ALE2_10_3, ALE5_2_9, and ALE_5_10_4 transformed with pCYS*_Cg(T94A)-SER* was lower than that of the aecD gene disruptant transformed with the same plasmid (Fig. 5a). The cysteine production levels in the pCYS*_Cg(T94A)-SER*-transformed cysteine-resistant mutants were higher than those in the pCYS*_Cg(T94A)-SER*-transformed aecD gene disruptant (Fig. 5b). This indicates that cysteine resistance is an effective phenotype for cysteine production by C. glutamicum.
Fig. 5
Cysteine production by the engineered cysteine-resistant mutants of C. glutamicum. Time courses of cell growth (a) and cysteine production (b) in the aecD disruptant (black circles), ALE2_10_3 (red triangles), ALE5_2_9 (blue squares), and ALE5_10_4 (green diamonds) transformed with pCYS*_Cg(T94A)-SER* are shown. For cultivation, a semisynthetic cysteine production medium was used. The average ± standard deviation of the data in three independent cultures is shown
Comments (0)