
Remember me
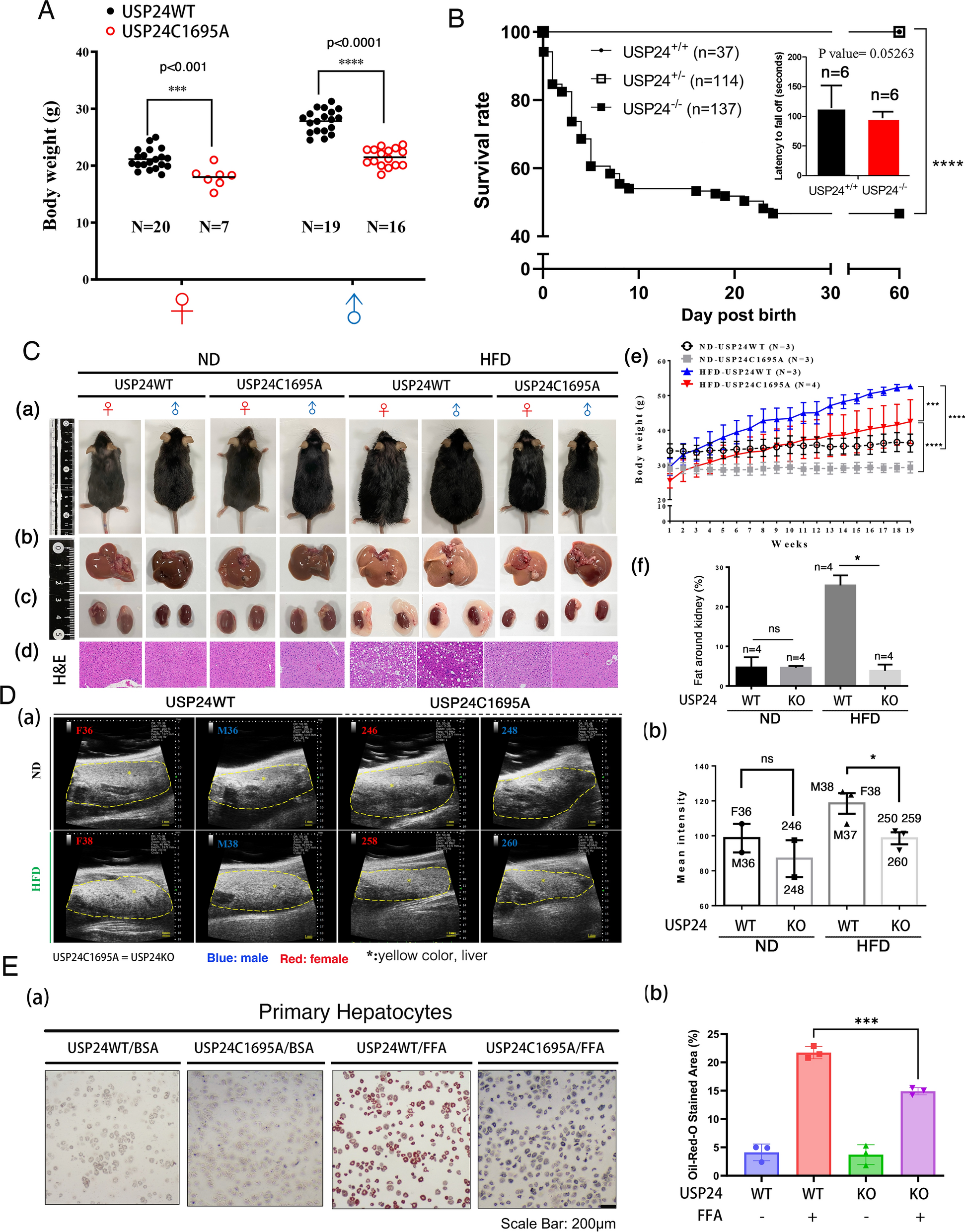


Previous studies on the role of USP24 in neurodegenerative diseases and cancer progression have been conducted, but its role in MAFLD progression remains unclear. According to previous studies on USP24, the loss of cys1698 (cys1695 in mice) in the catalytic motif of USP24 results in enzyme-death [30]. Therefore, we used CRISPR/Cas9 to construct mice with functional knockout of USP24 (USP24C1695A). The added Nar I restriction enzyme site was used for genotyping, and the experimental design and genotyping results are shown in Suppl. Figure 1A and Suppl. Figure 1B. The body weights and sizes of USP24C1695A (USP24KO) mice were lower than those of USP24WT mice, implying that USP24 expression might be involved in lipogenesis (Fig. 1A and Suppl. Figure 1C). There was no significant difference in food intake between USP24WT and USP24C1695A mice (Suppl. Figure 1D). The USP24+/+ and USP24± mice survived, but approximately 50% of the USP24−/− (USP24C1695A/C1695A) newborn mice died 1 week after birth (Fig. 1B). However, the mice who survived the first week after birth tended to survive, and the physical activity of the USP24WT and USP24C1695A mice was not significantly different (Fig. 1B, right panel). USP24WT and USP24C1695A (USP24−/−) mice fed a high-fat diet (HFD) for 19 weeks were used to study the role of USP24 in lipogenesis (Fig. 1C and D). Compared with USP24WT mice, USP24C1695A mice had significantly decreased body weights under normal diet (ND) and high-fat-diet (HFD) conditions (Fig. 1C, e). The blood glucose levels in USP24C1695A mice were lower than those in USP24WT mice under HFD conditions (Suppl. Figure 1E). The lipid droplet formation (Suppl. Figure 2A) and insulin expression (Suppl. Figure 2B) were decreased, and there was no significant change in pancreatic β-cells in USP24C1695A mice (Suppl. Figure 2C), implying that USP24 might also be involved in diabetes (Suppl. Figure 2). After 19 weeks of ND or HFD feeding, USP24WT HFD-fed mice were larger than USP24C1695A HFD-fed mice were, suggesting that the loss of USP24 enzyme activity inhibited obesity (Fig. 1C, a). Furthermore, lipid droplets accumulated in the livers of USP24WT HFD-fed mice but not in those of USP24C1695A HFD-fed mice, indicating that the loss of USP24 may inhibit fatty liver development (Fig. 1C, b, d). Interestingly, visceral adipose tissue (VAT) around the kidneys was found in HFD-USP24WT-mice but not in ND- and HFD-USP24C1695A-mice (Fig. 1C, c, f), indicating that the loss of USP24 enzyme activity inhibits fat accumulation in vivo. Ultrasound was also used to study the fat accumulation in vivo (Fig. 1D). The data revealed an obvious signal in the livers of HFD-USP24WT male and female mice but not in those of HFD-USP24C1695A mice (Fig. 1D, a, b and Suppl. Figure 1F). The role of USP24 in lipogenesis in hepatocytes was investigated (Fig. 1E and Suppl. Figure 3). First, we used a hepatoma cell line, Huh-7 cells, to study this question (Suppl. Figure 3). USP24-i-101, which targets USP24, did not inhibit lipogenesis in Huh-7 (Suppl. Figure 3A-3C). Furthermore, we directly cultured primary hepatocytes from USP24WT and USP24C1695A mice to study the effect of USP24 expression on hepatocyte lipogenesis (Fig. 1E). Compared with those from USP24WT mice, primary hepatocytes cultured from the livers of USP24C1695A mice showed dramatically decreased FFA-induced lipid droplet formation (Fig. 1E, b). In summary, USP24 expression not only facilitates fatty liver but also facilitates the accumulation of visceral adipose tissue (VAT) around organs and subcutaneous adipose tissue (SAT), which induces several related diseases, such as liver cirrhosis, cardiovascular disease, and diabetes.
Fig. 1
Functional knockout of USP24 suppresses lipogenesis. A Body weights of USP24WT and USP24C1695A mice fed a normal diet were measured. B Survival rate of newborn mice (B, left panel) and physiological activities of mice (B, right panel) with or without USP24 knockout were assessed by tracing the survival rate and performing a rotarod test, respectively. The results of A and B were analyzed by statistical assay, t-test, ***p < 0.001, ****p < 0.0001. USP24WT (n = 3) and USP24C1695A (n = 3) mice and USP24WT (n = 3) and USP24C1695A (n = 4) mice were fed a normal diet (ND, 13.1% fat) or a high-fat-diet (HFD, 60% fat) for 4 months, respectively. Mouse body sizes (C, a), organs (liver and kidney) (C, b, c), and H&E staining (C, d) are shown. The body weight of mice (C, e) and fat around kidneys (C, f) were quantified, and statistical analysis was performed using a t-test, *p < 0.05, ***p < 0.001, ****p < 0.0001. Fat content inside ND- and HFD-fed mice with or without function knockout mice (USP24WT & USP24C1695A) was determined using an ultrasound machine (D, a), and the signal highlight with yellow dotted circle was quantified by Image J, and statistical analysis was conducted by t-test, *p < 0.05 (D, b). Lipid droplet formation in primary hepatocytes isolated from USP24WT and USP24C1695A mice treated with free fatty acid (500 μM FFA) was examined via oil red O staining assay (E, a). Subsequent quantification of lipid signaling levels followed three independent experiments, with statistical analysis performed using a t-test, ***p < 0.001 (E, b)
USP24-i-101, which targets USP24, reduces the size of adipose tissuesWe employed a specific USP24 inhibitor, USP24-i-101, to assess the therapeutic effect of USP24-i-101 on inhibiting lipogenesis mediated by USP24 (Fig. 2). Mice were pre-fed a HFD for 2 months and subsequently treated with various doses of USP24-i-101 twice a week for one month (Fig. 2A). The food intake of HFD-fed mice was not different from that of control mice with or without USP24-i-101 treatment (Suppl. Figure 4A). The body weights of the mice were measured every week (Fig. 2B). Compared to DMSO-treated mice, the body weights of USP24-i-101-treated mice were decreased by approximately 3–5 g (Fig. 2B). The body, VAT and SAT sizes of the mice were significantly decreased following USP24-i-101 treatment (Fig. 2D and Suppl. Figure 4B). Subcutaneous and visceral fat contents were measured via MRI (Fig. 2D). The data showed that the MRI signals representing subcutaneous adipose tissue (SAT) fat (Fig. 2D, a, b) and visceral adipose tissue (VAT) were increased in HFD-fed female and HFD-fed male mice, but 10 mg/kg USP24-i-101 in HFD-fed female mice and 1 mg/kg USP24-i-101 in HFD-fed male mice significantly inhibited fat signaling (Fig. 2D). After sacrifice, the fat collected from different locations was compared between HFD-fed mice that had or had not received USP24-i-101 treatment (Fig. 2E, a, b). The data indicated that visceral fat surrounded the kidneys in HFD-fed mice but was nearly abolished under USP24-i-101 treatment, suggesting that targeting USP24 dramatically inhibited obesity. The sizes of adipocytes in epididymal white adipose tissue (eWAT) and inguinal white adipose tissue (iWAT) were also studied (Fig. 2F). The sizes of adipocytes were increased in HFD-fed mice but were reversed after USP24-i-101 treatment (Fig. 2F). We also cultured wild-type primary hepatocytes to treat them with USP24-i-101 and found that USP24 expression significantly inhibited lipid droplet formation (Fig. 2G and Suppl. Figure 4C). In addition, serums collected from mice were used to investigate the biochemical analysis, and results indicated that total protein (TP), albumin (ALB) and triglyceride (TG) levels were not significantly different. GPT/GOT and TCHO expression were markedly inhibited under USP24-i-101 treatment, suggesting that USP24-i-101 is safe at the working dose (Fig. 2H).
Fig. 2
USP24-i-101, which targets USP24, inhibits lipogenesis. USP24WT and USP24C1695A mice were fed a normal diet (ND; n = 6) or a high-fat diet (HFD; n = 33) for 2 months and then treated with 0.5 mg/kg (n = 6), 1.0 mg/kg (n = 6), 5 mg/kg (n = 6), 10 mg/kg (n = 4) or 20 mg/kg (n = 5) USP24-i-101 twice a week for 1 month (A). Body weight changes in all mice were measured weekly (B). Mouse sizes and their lipid, SAT and VAT, contents, are presented (C). Lipid signals in USP24WT and USP24C1695A mice on either an ND or HFD with or without USP24-i-101 treatment were measured using MRI (D, a). The signal was quantified by Image J, and statistical analysis was performed by t-test, ***p < 0.001, ****p < 0.0005, ns: non-significant (D, b). Visceral adipose tissue (VAT) around the kidneys of female and male mice with or without USP24-i-101 treatment was shown (E, a), The area of VAT was quantified by Image J, and statistical analysis was conducted by t-test, ***p < 0.001 (E, b). iWAT and eWAT in male mice on either an ND or HFD with or without 1 mg/kg USP24-i-101 treatment are depicted (F, a). Adipocyte sizes were measured using Image J, and statistical analysis was performed by t-test, **p < 0.01, ***p < 0.001 (F, b). Primary hepatocytes isolated from the mice were treated with various doses of USP24-i-101 and FFA for 24 h, after which lipid droplet formation was studied by oil red O staining. After three independent experiments, lipid droplets contents were quantified, and statistical analysis was performed via a t-test, **p < 0.01 (G). Serum collected from the mice was used to study the levels of several lipogenesis-related markers, such as total protein (TP), GPT/ALT, GOT/AST, ALB, total cholesterol (TCHO) and triglyceride (TG) levels, via ELISA, followed by statistical analysis via a t-test, **p < 0.01, ***p < 0.001 (H)
USP24 promotes adipogenesisNext, we investigated the mechanisms through which USP24 regulates body weight and lipid accumulation. Previous studies have shown that adipocyte formation is a major factor in increased body weight and lipid accumulation. Here, we studied the effect of USP24 on adipocyte formation using oil red O staining (Fig. 3). 3T3-L1 cells were differentiated into adipocytes with or without USP24-i-101 treatment (0-8th day) (Fig. 1A). The data indicated that lipid droplets were dramatically accumulated in control differentiated cells (treated with DMSO) but could be inhibited after USP24-i-101 treatment, suggesting that USP24 expression positively regulates adipogenesis (Fig. 3A). To study the effect of USP24-i-101 on differentiation stage of adipocytes, USP24-i-101 was added at the early stage (0-2nd day) or late stage (4th–8th day) to study adipocyte differentiation. The data indicated that the addition of USP24-i-101 at an early stage inhibited adipocyte formation, while the addition of USP24-i-101 at a late stage did not, implying that USP24 expression is involved in adipocyte differentiation at an early stage (Fig. 3A). USP24 expression was also silenced during 3T3-L1 differentiation to address the role of USP24 in lipogenesis (Fig. 3B). According to cell morphology (Fig. 3B, a) and oil red O staining results (Fig. 3B, b, c), lipid droplet accumulation was nearly abolished in USP24-knockdown cells, indicating that USP24 expression is critical for lipogenesis in adipocytes. Finally, GFP, GFP-USP24 and GFP-USP24C1695A were overexpressed in 3T3-L1 cells to study adipogenesis (Fig. 3C). The data indicated that GFP-USP24 overexpression can significantly increase lipid droplet formation but can be inhibited by GFP-USP24C1695A overexpression, suggesting that the enzyme activity of USP24 is required for adipogenesis (Fig. 3C).
Fig. 3
Upregulation of USP24 promotes adipogenesis. The schedule for adipocyte differentiation from 3T3-L1 cells and the cellular morphology with or without USP24-i-101 treatment at different time points, days 0–2, days 0–8 and days 4–8, are shown (A, a, b). Lipid droplet content during adipogenesis with or without 10 μM USP24-i-101 treatment were measured via oil red O staining (A, c), and the number of lipid droplets (day 0–8) was quantitated using Image J (A, d). 3T3-L1 cells with or without USP24 expression knockdown by shUSP24 lentivirus were observed (B). The morphology of the cells was observed (B, a), and fat accumulation was determined by oil red O staining (B, b and c). The levels of fat in adipocytes were quantified after three independent experiments (B, d). GFP, GFP-USP24WT, and GFP-USP24.C1698A were overexpressed in 3T3-L1 cells during adipogenesis. Lipid droplet formation was measured by oil red O staining (C, a). After three independent experiments, the contents of lipid droplets were quantified via Image J (C, b). Statistical analysis was performed via a t-test, **p < 0.01, ***p < 0.001, ****p < 0.0001
USP24 increases p-CREB, CEBPβ and PPARγ expression to promote adipogenesisTo elucidate how USP24 regulates adipogenesis, 3T3-L1 cells were treated with USP24-i-101, which targets USP24, and the knockdown or overexpression of USP24 was investigated in differentiated 3T3-L1 cells (Fig. 4). The expression of USP24 increased during adipocyte differentiation, suggesting that USP24 expression might be related to adipocyte differentiation in the early period (Fig. 4A, a, b). The expression levels of several lipogenesis-related proteins, including SREBP1c, CREB, p-CREB, C/EBPβ,C/EBPδ, PPARγ, PLIN1, and LC3B, were determined in USP24-i-101-treated adipocytes (Fig. 4A, a, d–i). Data showed that the expression of all proteins except C/EBPδ was significantly inhibited by USP24-i-101 treatment (Fig. 4A, a, d–i). As LC3B expression was also increased here, USP24-i-101 was used to study its effect on autophagy in hepatocytes (Suppl. Figure 5). The data indicated that USP24-i-101 treatment increased LC3B expression in hepatocytes and adipocytes, suggesting that USP24-i-101-mediated autophagy might be involved in adipogenesis (Suppl. Figure 5A-5E). The expression of several lipogenesis-related proteins, SREBP1c, C/EBPβ, PPARγ, PLIN1 and p300, was determined in USP24-silenced adipocytes (Fig. 4B). Data indicated that the expression of all related proteins was significantly decreased in USP24-knockdown cells, implying that the increase in USP24 expression during 3T3-L1 differentiation is involved in lipogenic gene expression (Fig. 4B). GFP, GFP-USP24, or GFP-USP24C1695A was overexpressed in 3T3-L1 cells to study the expression of these adipogenesis-related proteins (Fig. 4C). Data indicated that the expression of all adipogenesis-related proteins, including p-SREBP1, PPARγ, C/EBPβ, PLIN1, CREB, and p-CREB, was increased in GFP-USP24-overexpressing cells but not in the GFP-USP24C1695A-overexpressing cells, indicating that the enzyme activity of USP24 is necessary for adipogenesis (Fig. 4C). Primary hepatocytes isolated from USP24 knockout mice were treated with free fatty acid (FFA), and FFA treatment significantly increased the expression of USP24 and PLIN2 in USP24WT mice but only slightly increased the expression of PLIN2 in USP24C1695A mice, suggesting that the enzyme activity of USP24 is important for FFA-mediated induction of lipogenesis in the liver (Fig. 4D). Inhibiting PKA activity with the PKA inhibitor H89 decreased USP24 expression (Fig. 4E), indicating that USP24 and PKA positively regulate each other. How does USP24 promote adipogenesis directly? The early-stage phosphorylation of CREB is critical for adipogenesis [31]. First, CREB protein phosphorylation during 3T3-L1 differentiation with or without USP24-i-101 treatment was measured (Fig. 4F). Data indicated that both the protein phosphorylation and total CREB but not mRNA expression level of CREB were decreased by USP24-i-101 treatment, implying that phosphorylation of CREB may be involved in its protein stability (Fig. 4F, Suppl. Figure 5G). Studying protein stability of p-CREB and CREB under cycloheximide (CHX) treatment found that phosphorylation of CREB can increase its protein (Fig. 4G), indicating that USP24 expression may increase the phosphorylation of CREB, thereby increasing its protein stability. These findings indicate that USP24 expression promotes adipogenesis through positive regulation of lipogenesis-related gene expression.
Fig. 4
The USP24/PKA/p-CREB/CEBPβ/PPARγ axis regulates adipocyte differentiation. Lipogenesis-related proteins expressed in 3T3-L1 cells treated with 10 μM USP24-i-101 (A) were knocked down with shRNA-USP24 (B) or overexpressed with GFP/GFP-USP24/GFP-USP24C1698A (C) during adipogenesis, and then samples were harvested on the -2nd, 0, 2nd, 4th, 6th and 8th days. The expression of lipogenesis-related proteins was measured via IB with antibodies agonist the indicated proteins (A–C). Primary hepatocytes isolated from mice with or without USP24 knockout, USP24WT or USP24.C1695A, were treated with 500 μM FFA, and then, the cells were lysed to study the expression of USP24 and PLIN2 via IB (D). 3T3-L1 cells were treated with MDIR and the PKA inhibitor 10 μM H89 for 24 h, and then the samples were collected in sample buffer. The expression of USP24 was measured via IB with anti-USP24 antibodies (E). 3T3-L1 cells with or without 10 μM USP24-i-101 treatment were lysed to study the expression of CREB and pCREB via IB (F, a). After three independent experiments, the expression of pCREB (F, b) and CREB (F, c) were quantified. 3T3-L1 cells were treated with cycloheximide (CHX), and then, cell lysates were collected at the indicated times to study the expression of pCREB/CREB (G, a). After three independent experiments, the expression of pCREB/CREB (D, b) and PKA-Cα (G, b) was quantified. After three independent experiments, various protein expressions were quantified. The statistical analysis was performed via a t-test, *p < 0.05, **p < 0.01, ***p < 0.001
USP24 expression stabilizes PKA-Cα and p300 during adipogenesisSince the data in Fig. 4 suggested that USP24 expression might be involved in the early period of adipogenesis, we studied the mechanism by which USP24 regulates signaling pathway during adipogenesis in detail (Fig. 5). Several kinases, including RSK2, CaMKII, CaMKIV and PKA, have been reported to phosphorylate CREB [31]. Our findings showed that treatment with USP24-i-101 or knockdown of USP24 expression decreased the expression of CaMKII and the PKA catalytic subunit α (PKA-Cα) (Fig. 5A and B). In contrast, overexpression of GFP-USP24WT in 3T3-L1 cells increased PKA-Cα expression compared with USP24C1698A overexpression (Fig. 5C, a, b). Additionally, the ubiquitination signal of PKA-Cα was reduced on day 4 compared to day 0 (Fig. 5C, c), suggesting that the enzyme activity of USP24 is required for adipogenesis. Treatment with USP24-i-101 and knockdown of USP24 expression also decreased the protein stability of PKA-Cα (Fig. 5D and E), implying that PKA-Cα may be as a substrate of USP24. Since p300 can bind to CREB to increase its activity [22], our previous studies revealed that p300 is the substrate of USP24 [31]. We also found that the knockdown of USP24 expression decreased the protein expression of p300 but did not significantly change its mRNA expression (Fig. 4B and Suppl. Figure 5F). PKA-Cα and p300 can interact with USP24 but not CaMKII (Fig. 5F). Overexpression of GFP-USP24 in 3T3-L1 cells increased PKA-Cα expression and decreased the ubiquitination signal of PKA-Cα through a proteasome-dependent manner, whereas this effect was not observed when GFP-USP24C1695A was overexpressed (Fig. 5G). Conversely, treatment with USP24-i-101 or knockdown of USP24 expression increased the ubiquitination signal of USP24 in a proteasome-dependent manner under similar PKA-Cα level condition (Fig. 5H, I). In in vitro enzyme assays revealed that USP24 can eliminate the ubiquitination of PKA-Cα, possibly including the polyubiquitination and monoubiquitination of PKA-Cα (Fig. 5J). Previous studies have indicated that two residues, K286 and K310, in PKA-Cα can be ubiquitinated by its E3-ligase [32]. A model of the USP24-ubiquitin-PKA-Cα complex was generated to gain better insight into its structure (Fig. 5K). The USP24 catalytic site is positioned close to the ubiquitin-PKA-Cα protein. Molecular Dynamic dynamics simulations revealed that USP24 residues are positioned near the USP24-ubiquitin-PKA-Cα complex, including USP24 residues C10, H282, and N302, which are positioned near the PKA-Cα G76 and ubiquitin K310 bonds. The position of USP24 facilitates tagging for additional cellular processing of the complex. Finally, a cAMP analog, rp-cAMPs, was employed to compete with cAMP, thereby inhibiting the activity of PKA-Cα, to address the synthesis of fatty acid in adipocytes (Suppl. Figure 5H). The synthesis of fatty acid was partially inhibited by rp-cAMPs, but can be totally inhibited by USP24-i-101, indicating that other USP24-regulated factors, such as CaMKII, also contribute to the effect of USP24 in adipogenesis (Suppl. Figure 5H). In summary, USP24 not only interacts with PKA-Cα to positively regulate each other, increasing CREB1 phosphorylation but also stabilizes p300 and increases the transcriptional activity of p-CREB during adipogenesis.
Fig. 5
USP24 stabilizes PKA-Cα and p300 during adipogenesis. The expression of USP24 in 3T3-L1 cells was knocked down using shRNA-USP24 for three days, after which the cells were differentiated into adipocytes. Samples were collected at different time points to study the expression of the indicated proteins via IB with antibodies against the indicated proteins (A, a). After three independent experiments, the expression of CaMKII (A, b) and PKA-Cα (A, c) was quantified. 3T3-L1 cells were treated with or without 10 μM USP24-i-101, and samples were collected at day 2 and day 4 (B, a) to measure the expression of PKA-Cα and CaMKII via IB. After three independent experiments, the expression of PKA-Cα (B, b) and CaMKII (B, c) was quantified. GFP-USP24 and GFP-USP24C1698A were overexpressed in 3T3-L1 cells, which were then induced for adipocyte differentiation via MDIR. Samples were collected at different time points to measure the expression of PKA-Cα via IB with anti-PKA-Cα antibodies (C). 3T3-L1 cells were treated with cycloheximide (CHX) with or without 10 μM USP24-i-101 treatment (D) and USP24 knockdown (E), and then cell lysates were collected at the indicated times to study the expression of PKA-Cα (D and E). After three independent experiments, the expression of PKA-Cα (D and E, b) was quantified. 3T3-L1 cells were differentiated, and samples were collected on the 2nd day for IP with anti-USP24 and IgG (F, a, c) or anti-PKA-Cα and IgG (F, b), and then IB with antibodies against indicated proteins. GFP, GFP-USP24WT and GFP-USP24C1698A were transfected into 3T3-L1 cells for differentiation, and samples were for IP with anti-GFP antibodies or anti-USP24 antibodies and IB with antibodies against USP24, PKA-Cα, ubiquitin and actin (G). 3T3-L1 cells with or without USP24-i-101 (H), or USP24 knockdown (I) were differentiated with MG132 treatment, and samples were collected on the 2nd day for IP with anti-ubiquitin and IgG and IB with antibodies against PKA-Cα. GFP and GFP-PKA-Cα were overexpressed in 3T3-L1 cells for 2 days. Polyubiquitinated PKA-Cα was pulldown by IP with anti-ubiquitin antibodies (J, a), anti-PKA-Cα and anti-GFP antibodies (J, b), followed by the addition of myc-USP24 (1 μg recombinant protein) for 1 h. The levels of various proteins were measured by IB with indicated antibodies (J). The generation of the USP24-ubiquitin-PKA-Cα complex may indicate its function. An irreversible bond is formed between ubiquitin G76 and PKA-Cα K310. The catalytic site of USP24 envelops the ubiquitin-PKA-Cα bond and can be near the USP24 catalytic residues C10, H282, and N302. The modelled complex suggests potential functions for further cellular processing of the complex (K)
USP24 positively regulates lipogenesis, inflammation, and fibrosis gene expression in HFD-fed miceTo study the effect of USP24 on gene expression systemically, liver organs collected from normal diet (ND)-fed and high-fat diet (HFD)-fed mice with or without USP24 functional knockout (USP24C1695A) were used to investigate the gene expression profiles via RNA-seq (Fig. 6, Suppl. Figure 6-Suppl. Figure 11 and Suppl. Table 1-Table 2). First, the concentration and quality of all the RNA samples were evaluated (Suppl. Figure 7A). All the control results, including the error rate distribution, base content distribution, Trimmomatic results, distribution of gene expression levels, Pearson correlation coefficient, Trimmomatic Trim results summary and principal component analysis (PCA), supported the quality of these RNA-seq results (Supp. Figure 6). After analysis with bioinformatics tools, 236 genes in the livers of USP24WT mice but only 52 genes in the livers of USP24C1695A mice were found to show upregulated expression in HFD-fed mice compared with ND-fed mice (Suppl. Figure 7B, b, c), implying that USP24 regulates lipogenesis-related gene expression. However, 103 genes in female USP24-knockout HFD-fed mice and 113 genes in male USP24C1695A HFD-fed mice showed significantly downregulated expression compared with those in USP24WT HFD-fed mice (Suppl. Figure 7B–Suppl. Figure 7C). Most expression of the genes related to fatty liver and diabetes, such as LPIN1, LCN2 and CTSB, in USP24C1695A HFD-fed mice were downregulated, suggesting that USP24 is involved in lipogenesis (Suppl. Figure 8). Acetyl-CoA carboxylase (ACACB), which catalyzes the carboxylation of acetyl-CoA to malonyl-CoA, the rate-limiting step in fatty acid synthesis, and ACACB expression was also significantly downregulated in USP24C1695A HFD-fed mice (Suppl. Figure 8). The abundance of stearoyl-CoA 9-desaturase 1 (SCD1), an iron-containing enzyme that catalyzes a rate-limiting step in the synthesis of unsaturated fatty acids, was dramatically decreased in USP24C1695A HFD-fed mice (Suppl. Figure 8). Another gene, secreted phosphoprotein 1 (SPP1), which is overexpressed in many cancer types and correlated with poor prognosis, was also significantly downregulated expression in USP24C1695A HFD-fed mice (Suppl. Figure 8) 30,31. By using GO enrichment (Suppl. Figure 9A) and DEG analysis, we also found that most of the genes related to lipogenesis, such as those related to fatty acid metabolism, acyl-CoA metabolism, steroid metabolism, and lipid catabolism, had downregulated expression in USP24-knockout HFD-fed mice (Fig. 6A, b and Suppl. Figure 9A). In addition, biological pathways (DEG Down GO TERMs-BP Enrichment; Fig. 6A, a and Suppl. Figure 10A) and disease pathways (DEG Down DisGeNET Enrichment Pathway; Suppl. Figure 9B and Suppl. Figure 10B) were regulated by USP24. Other genes related to diseases such as lupus vulgaris, amyloid neuropathies, aortic aneurysm and chronic kidney diseases also showed downregulated expression in USP24C1695A mice (Suppl. Figure 10B). Many lipogenesis-related diseases, such as fatty liver, hyperinsulinism, diabetes, and dyslipidemia, were significantly inhibited in USP24C1695A -overexpressed or USP24-i-101-treated HFD-fed mice, implying that targeting USP24 might prevent these diseases (p < 0.00005; Fig. 6A, a and c). In the DEG Down GO TERM-CC enrichment analysis, we found that the loss of USP24 expression decreased the activity of several lipid trafficking pathways, such as the lysosome, lytic vacuole and lipoprotein particle pathways (Suppl. Figure 10C). In the DEG downregulated KEGG enrichment analysis, pathways related to lysosomal activity, biosynthesis of unsaturated fatty acids, the PPAR signaling pathway and alcoholic liver disease had downregulated expression in USP24-knockout HFD-fed mice (Suppl. Figure 11A). In the DEG Down DO Enrichment Pathway, pathways related to arteriosclerotic cardiovascular disease obesity were significantly inhibited by knocking out USP24 (Suppl. Figure 11B). Numerous genes related to β-oxidation (Suppl. Figure 12A, a), adipogenesis (Suppl. Figure 12B, b), glycolysis and gluconeogenesis (Suppl. Figure 12C, a, b) had downregulated expression in USP24C1695A mice (Suppl. Figure 12).
Fig. 6

Numerous genes related to lipogenesis are regulated by USP24. Total RNA samples were isolated from the liver tissues of USP24WT, USP24C1695A or USP24-i-101-treated mice fed a ND or HFD for RNA-Seq and then analyzed via the bioinformatics tools DEG Down GO TERMs – BP Enrichment (A, a; USP24C1695A/USP24WT), DEG All Top 20 GO Barplot (A, b; USP24C1695A/USP24WT), DEG Down ALL GO Dotplot (A, c; 1 mg/KG USP24-i-101, i.p., two times per week) and Volcano Plot (A, d; 1 mg/KG USP24-i-101, i.p., two times per week). Inflammation-related genes whose expression was regulated by USP24C1695A were studied via RNA-seq and analyzed with a heatmap (B, USP24C1695A/USP24WT). Total RNA was extracted from mice with or without USP24 expression knockout to study the mRNA expression of Ly86 (C, a) and pLa2g7 (C, b) via qPCR. AML12 cells with or without USP24 knockdown were treated with LPS for 2 h, and the mRNA expression of USP24 (D, a) and Pla2g7 (D, b) were measured via qPCR. Primary hepatocytes isolated from mice with or without USP24 knockout (E, a, b) or hepatocyte AML cells (E, c, d, e) were treated with LPS or USP24-i-101. The expression of USP24, p-NF-κB and NF-κB were measured via IB (E). AML12 cells with or without USP24 knockdown (F) or 10 μM USP24-i-101 treatment (G) were treated with 500 μM free fatty acid (FFA) or BSA for 24 h, after which the samples were harvested to measure the expression of USP24,COX2, IκB, p-NF-κB and NF-κB via IB. After three independent experiments, the quantification and statistical analysis were performed via t-test: *p < 0.05, **p < 0.01, ***p < 0.001
We examined the global gene expression profiles not only in HFD-USP24C1695A mice but also in USP24-i-101-treated mice (Fig. 6A, c and d, Suppl. Figure 9C-9E). All control results, including H&E staining results of the RNA-seq samples (Suppl. Figure 13), the error rate distribution (Suppl. Figure 14A), the base content distribution (Suppl. Figure 14B), Trimmomatic results (Suppl. Figure 14C), the distribution of gene expression levels (Suppl. Figure 14D), Pearson correlation coefficient (Suppl. Figure 14E) and Trimmomatic Trim results summary (Suppl. Figure 14F), supported the quality of these RNA-seq results. We used a heatmap (Suppl. Figure 9C) and volcano plot (Fig. 6A, d) to address the effect of USP24-i-101, which targets USP24, on the systemic gene expression profile of HFD-fed mice (n = 3). A total of 332 genes showed downregulated expression, and 394 genes showed upregulated expression following the 5 μM USP24-i-101 treatment (Suppl. Figure 9C and Fig. 6A, d). Several lipocalin (LCN)-like genes, such as Mup1, Mup3, Mup12, Mup15, Mup16, Mup7, Mup11, Mup17 and Mup14, had upregulated expression induced by USP24-i-101 treatment, which is consistent with the inhibition of obesity and diabetes [33, 34]. Several Cyp genes, such as Cyp4a14, Cyp3a11 and Cyp4a10, had downregulated expression in mice treated with USP24-i-101, which targets USP24 (Fig. 6A, d). Several lipogenesis-related pathways, such as the fatty acid metabolic process, lipid catabolic process, alcohol metabolic process, cholesterol metabolic process and steroid metabolic process, were downregulated in DEG ALL GO Dotplot (Suppl. Figure 9D) and DEG Down ALL GO Dotplot (Fig. 6A, c). However, several pathways related to the collagen-containing extracellular matrix, negative regulation of proteolysis and the humoral immune response had decreased activity according to the DEG Up ALL GO Dotplot (Suppl. Figure 9E). Finally, not only fatty acid metabolism-related genes (Suppl. Table 1) but also inflammation-related genes (Fig. 6B and Suppl. Table 2) and fibrosis-related genes (Suppl. Table 3) had downregulated expression in USP24C1695A mice, implying that targeting USP24 might potentially inhibit metabolic associated fatty liver disease (MAFLD). The expression of two inflammatory-related genes, Ly86 and Pla2g7, was validated in mice with or without USP24 knockout by qPCR (Fig. 6C). Indeed, the loss of USP24 expression decreased the mRNA expression of Ly86 and Pla2g7 (Fig. 6C). Hepatocyte AML12 cells with or without USP24 knockdown were treated with LPS to measure the expression of Pla2g7 (Fig. 6D). The data indicated that USP24 knockdown decreased the mRNA expression of LPS-induced Pla2g7 in AML12 cells (Fig. 6D). WT and USP24-knockout hepatocytes isolated from mice (Fig. 6E, a, b) or hepatocyte AML cells (Fig. 6E, c, d, e) were used to study the effect of USP24 expression on inflammation with or without LPS or USP24-i-101 treatment. The levels of NF-κB and p-NF-κB were significantly decreased in USP24C1695A and USP24-i-101-treated USP24WT primary hepatocytes (Fig. 6E, a, b) and AML cells (Fig. 6E, c, d, e), suggesting that the loss of USP24 expression in mice decreases the activity of the NF-κB signaling pathway, thereby inhibiting inflammation (Fig. 6E). Finally, the effect of USP24 expression on inflammation in FFA-treated AML cells with or without USP24 knockdown (Fig. 6F) or USP24-i-101 treatment (Fig. 6G) was also investigated. The expressions of NF-κB and p-NF-κB (p-p65) were inhibited in USP24-knockdown (Fig. 6F) or USP24-i-101-treated (Fig. 6G) FFA-treated AML cells. Together, these findings suggest that USP24 promotes inflammation and fibrosis during MASH progression. In summary, not only in USP24 knockout mice but also in USP24-i-101-treated mice, most lipogenesis-, inflammation- and fibrosis-related genes had downregulated expression, indicating that USP24-i-101, which targets USP24, can inhibit MASH.
Inhibition of USP24 expression suppresses inflammation- and fibrosis-related gene expressionTo date, we have elucidated the role and mechanism of USP24 in adipocytes. Does USP24 expression participate in MASH progression in the liver? To further investigate the role of USP24 expression in the liver, several experiments related to MASH were conducted here (Fig. 7). All the pathways associated with fibrosis showed decreased activity in USP24 knockout mice (Suppl. Figure 15A). The number of cells with macrovascular steatosis in the livers of the mice was determined (Suppl. Figure 14 and Suppl. Figure 15B). The number of cells with macrovascular steatosis was higher in HFD-fed mice than in ND-fed mice, and the knockout of USP24WT (USP24C1695A) decreased the number of these cells (Suppl. Figure 15B and Suppl. Figure 16). Additionally, the mRNA expression of fibrosis-related genes α-SMA, Cola1 and TGF-β1 was significantly inhibited by USP24-i-101 treatment in primary hepatocytes (Suppl. Figure 15C). USP24 expression was knocked down or treated with USP24-i-101 in LX-2 cells (Fig. 7A and B) and AML12 cells (Fig. 7C and D) with or without LPS treatment to study the expression of inflammation- and fibrosis-related proteins. The expression of COX2, NF-κB, p-p65, vimentin, NLRP3, N-cadherin (N-cad), α-SMA, fibronectin and collagen were significantly decreased, and IκB expression was increased in LX-2 cells (Fig. 7A and B) and AML12 cells (Fig. 7C and D), suggesting that USP24 expression positively regulates inflammation in the liver. Since TGF-β can induce fibrosis in the liver, USP24 plays a role in TGF-β-treated LX-2 cells (Fig. 7E and F) and AML12 cells (Fig. 7G and H). The expression of the fibrosis-related proteins vimentin, N-cadherin, α-SMA, fibronectin and collagen were inhibited in USP24-knockdown LX-2 cells (Fig. 7E) and AML cells (Fig. 7G) and in USP24-i-101-treated LX-2 cells (Fig. 7F) and AML cells (Fig.
Comments (0)