
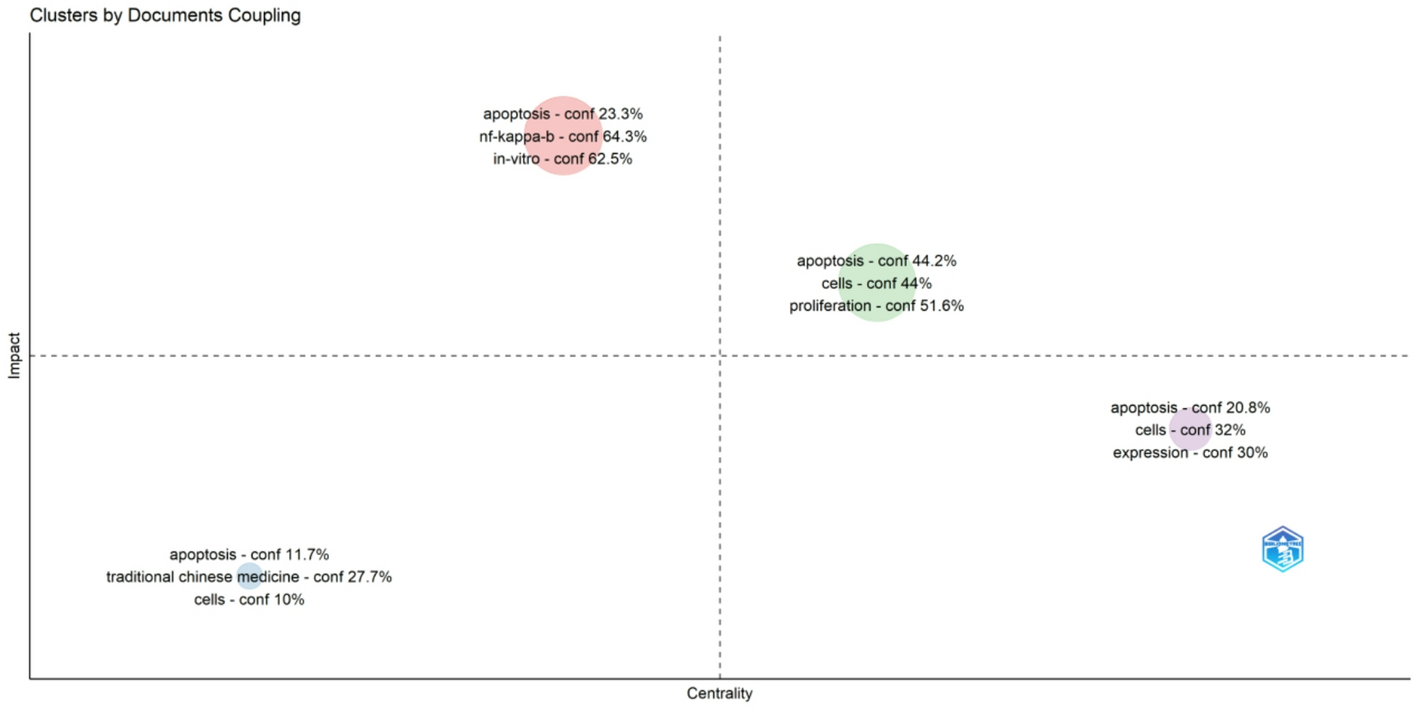
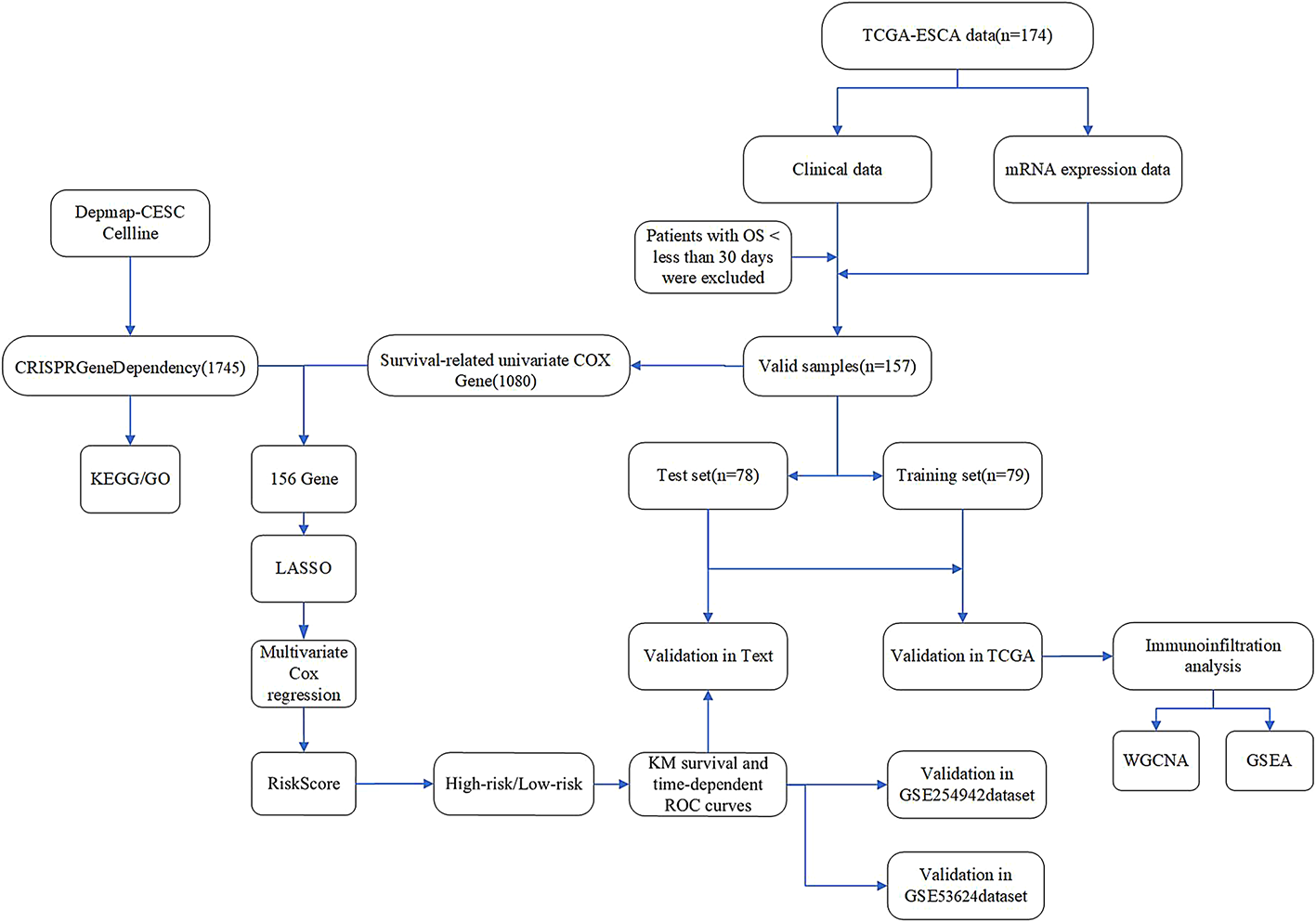
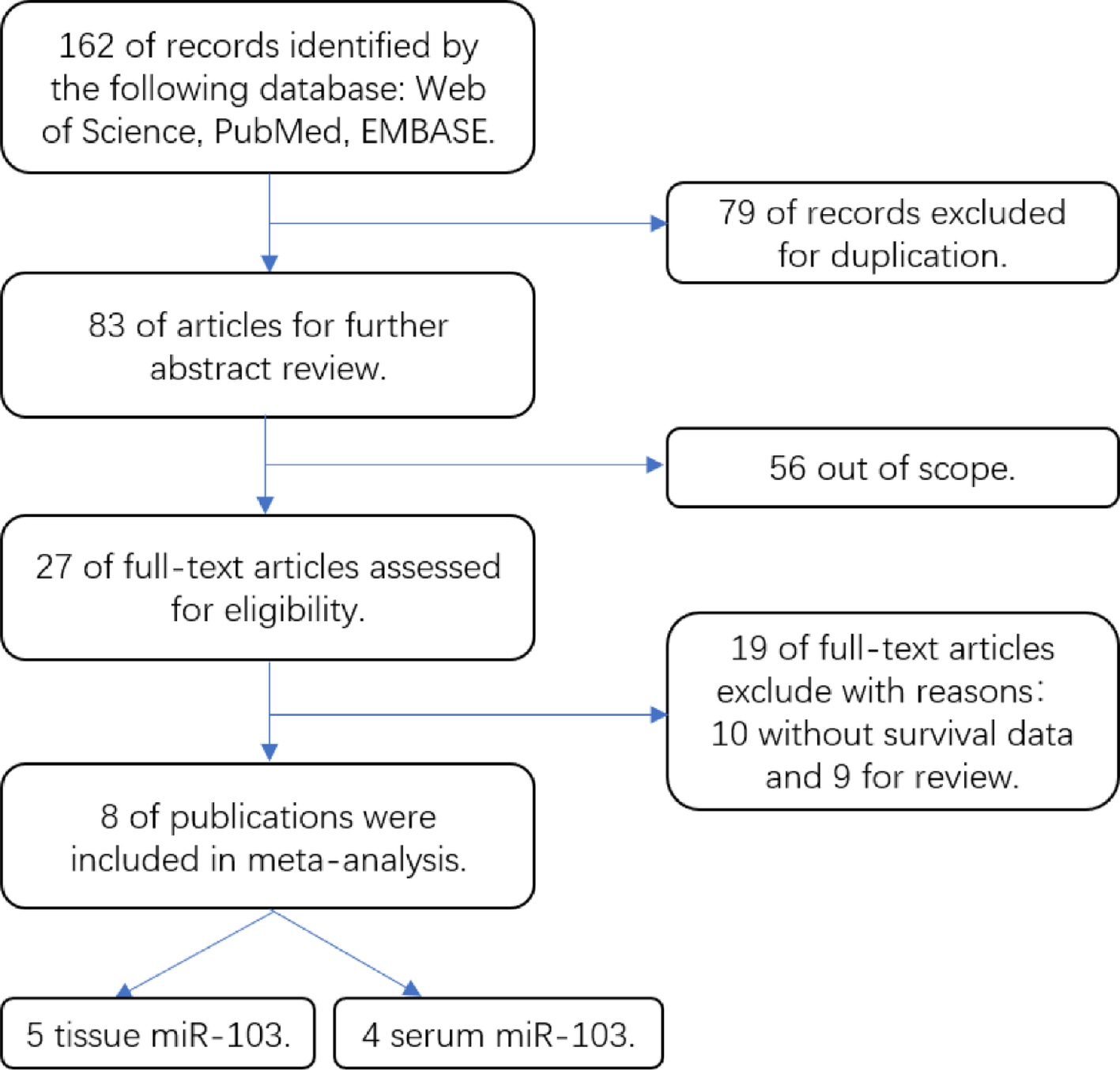
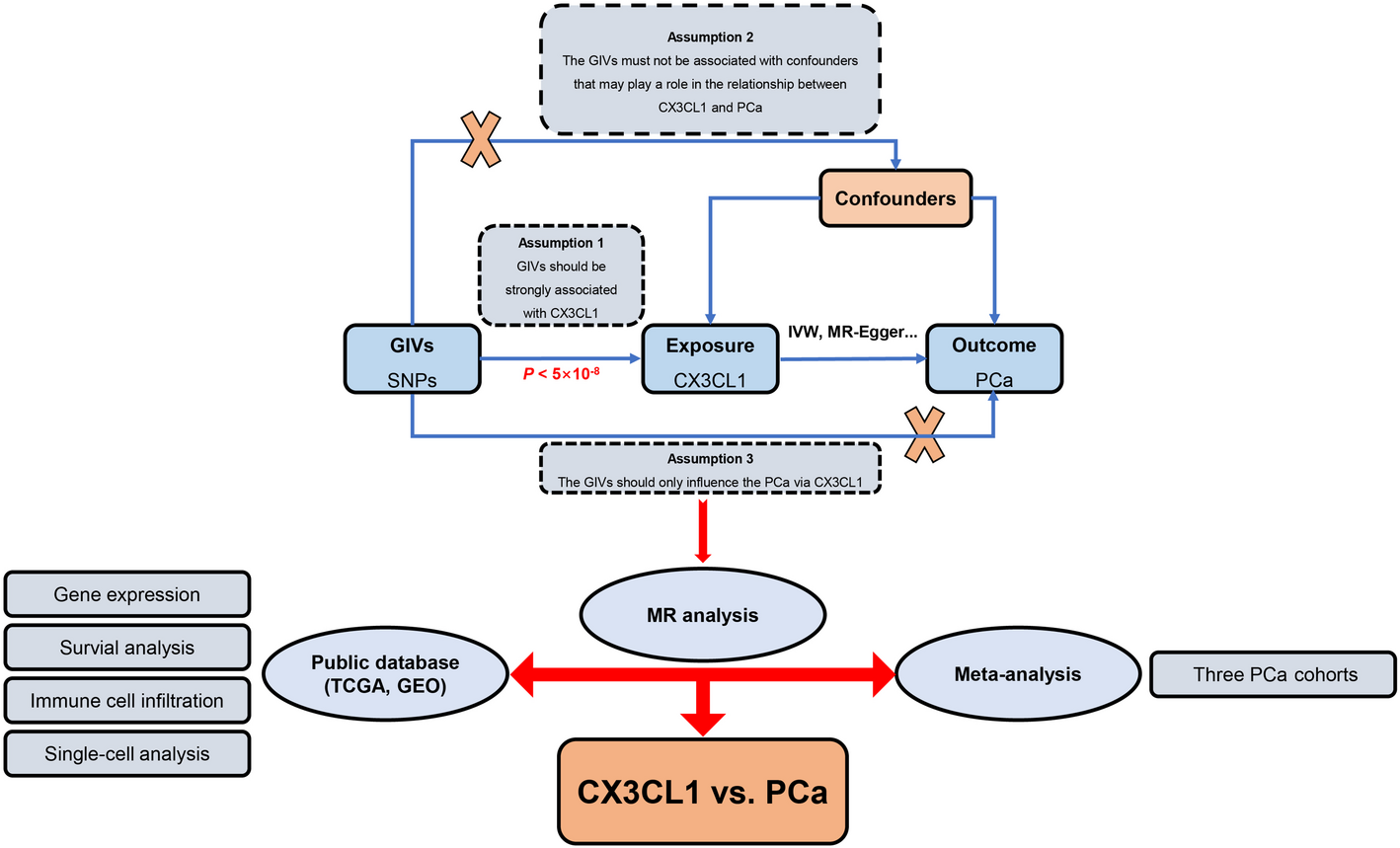
Remember me


Cervical cancer target genes were identified using GeneCards and DisGeNET databases, yielding 7846 putative target genes related to cervical cancer. Furthermore, targets related to CQ were extracted using the PharmMapper, SwissTargetPrediction, and Drug databases, yielding 126 drug targets. Of the 7846 possible oncogene targets related to cervical cancer, 126 CQ targets intersected with 100 targets (Fig. 2, Supplementary file 1), which were considered candidate targets.
Fig. 2
VENN plot of chloroquine targets versus cervical cancer targets
3.1.2 Construction of PPI network and key target screeningAfter identifying potential target genes of CQ in cervical cancer using the STRING website, a gene–gene interaction network comprising 93 nodes and 640 edges was constructed (Fig. 3). Among these, TNF, MDM2, SLC6A4, GRIN1, CHRM1, EZH2, ACHE, and ADRB2 were identified as the core genes with the highest degree of centrality in the PPI network, with a significant number of edges connecting these nodes (37, 16, 21, 14, 14, 15 12, 12, 10, and 18, respectively).
Fig. 3
Protein–protein interaction (PPI) network map of the targets common to both chloroquine and cervical cancer. a Visualisation of the target proteins in the STRING database. b PPI network obtained from further data processing using Cytoscape. This network represents potential therapeutic targets for chloroquine against cervical cancer. The larger the node, the higher the corresponding target degree and the greater the number of connections with other nodes
3.1.3 GO and KEGG enrichment analysesGO enrichment analysis, including biological process (BPs), cell composition (CCs), and molecular function (MFs), was performed to further analyse the identified candidate target genes. In terms of BPs, the genes were mainly enriched in ‘response to xenobiotic stimulus’, ‘response to the drug’, ‘intracellular signal transduction’, and ‘positive regulation of cell proliferation’. In terms of CC, the genes were primarily enriched in ‘plasma membrane’, ‘integral component of plasma membrane’ and ‘synapse’. Most of the genes in the MF group are enriched in ‘protein binding’, ‘protein kinase binding’, and ‘enzyme binding’. KEGG pathway enrichment was used to investigate the primary role of CQ, revealing the potential involvement of 30 signalling pathways (Fig. 4) (P < 0.05). The main pathways associated with cervical cancer included the PI3K/AKT, cAMP, neuroactive ligand-receptor interaction, C-type lectin receptor, and VEGF signalling pathways.
Fig. 4
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genome (KEGG) enrichment analysis. a Gene Ontology enrichment analysis. BP, biological process; CC, cell composition; MF, molecular function; b KEGG enrichment analysis of therapeutic targets
3.2 In vitro experiment validation3.2.1 CQ inhibits the proliferation of HeLa cellsThe CCK-8 assay was used to investigate the effect of CQ on the proliferation of HeLa cells and H8 cells. For 24 and 48 h, HeLa cells and H8 cells were treated with various doses of CQ (25, 50, 75, 100, and 150 µM). CQ significantly suppressed HeLa cell proliferation in a time-dependent manner (Fig. 5a) (P < 0.05). At 48 h, 75 µM CQ decreased HeLa cell growth by 50%. The proliferation of H8 cells was not obviously changed with the increasing concentrations(25µM, 50µM, 75µM) of CQ at each time point (Fig. 5b).However, the proliferation of H8 cells was decreased with the increasing concentrations of CQ (100, and 150 µM). At identical drug concentrations and treatment durations, CQ demonstrated significantly higher cytotoxicity against HeLa cells than against H8 cells.As shown in(Fig. 5c), the clone formation of the HeLa cells was suppressed with 25 µM and 75 µM of CQ treatment compared with no suppression in the control group.
Fig. 5
Effect of chloroquine on HeLa cell survival. a CCK-8 assay of the survival of HeLa cells treated with different concentrations of CQ for 24 and 48 h.bCCK-8 assay of the survival of H8 cells treated with different concentrations of CQ for 24 and 48 h. c Clone formation assay of HeLa cells treated with different concentrations of CQ for 48 h. *P < 0.05, **P < 0.01,***P < 0.001
3.2.2 CQ prevents HeLa cells from forming coloniesThe effect of CQ on colony formation by HeLa cells was investigated using a colony formation assay. The results showed that the colony formation rate of the experimental group was much lower than that of the control group, and colony formation declined progressively as the CQ concentration increased (Fig. 5b). A quantitative examination of colony formation by HeLa cells at various concentrations of CQ (25 and 75 µM) revealed a significant difference between the control and experimental groups (P < 0.05).
3.2.3 CQ inhibits the migration of HeLa cellsA scratch assay was performed to investigate the effect of CQ on the migratory capacity of HeLa cells. In this experiment, the migratory capacity of cells in the experimental group was dramatically reduced in a dose-dependent manner compared to that in the control group, which did not receive CQ or received an equivalent volume of solvent without CQ (Fig. 6). Quantitative examination of the migration of HeLa cells treated with various doses of CQ (25 and 75 µM) revealed a significant difference between the control and experimental groups (P < 0.05).
Fig. 6
Effect of chloroquine on the cell migratory ability of HeLa. Migration of HeLa cells treated with different concentrations of CQ for 48 h; *P < 0.05,**P < 0.01, ***P < 0.001
3.2.4 CQ induces apoptosis in HeLa cellsTo investigate the impact of CQ on the apoptosis of HeLa cells, we analyzed the apoptosis rates of HeLa cells treated with 0, 25, and 75 µM CQ for 48 h. Apoptosis was detected using flow cytometry in HeLa cells, which were labelled with Annexin V-FITC and 7-AAD. The results demonstrated that the apoptosis rate in the experimental group was much higher than that in the control group, exhibiting a dose-dependent relationship (Fig. 7a) (P < 0.05). Further studies of the molecular mechanisms indicate that 75µM of chloroquine CQ significantly inhibits the PI3K/AKT signaling pathway, reduces the expression of anti-apoptotic proteins such as Bcl-2, and simultaneously increases the expression of pro-apoptotic proteins such as Bax. Next, we evaluated the expression of apoptotic markers in the presence of CQ. As the dose of CQ increased, the level of cleaved poly(ADP-ribose) polymerase (cleaved-PARP) in HeLa cells also increased, as demonstrated in(Fig. 7b) (P < 0.05).
Fig. 7
Effect of chloroquine on HeLa cell apoptosis. a Flow cytometry analysis of HeLa cells treated for 48 h. b Western blot analysis of the relative expression of apoptotic proteins (Bax/Bcl-2, and cleaved PARP) in HeLa cells treated for 48 h with different concentrations of CQ. *P < 0.05, **P < 0.01, ***P < 0.001
3.2.5 CQ inhibits the PI3K/AKT/MDM2 pathwayWith the purpose of investigating whether CQ is associated with the induction of apoptosis by blocking the PI3K/AKT/MDM2 pathway in the long term, we conducted UALCAN and Western-Blot analysis to examine the expression of appropriate proteins. As shown in (Fig. 8, Supplement Fig. 1) (P < 0.05), treatment with CQ significantly reduced the phosphorylation of PI3K, AKT. CQ can also inhibit the expression of p-MDM2. The results indicate that the mechanism of CQ is based on cell appropriate.
Fig. 8
Effect of chloroquine on the expression of the PI3K/AKT pathway proteins. Western blot analysis of the relative expression of p-PI3K, p-AKT, and p-MDM2 proteins in HeLa cells treated with different concentrations of CQ for 48 h. *P < 0.05, **P < 0.01, ***P < 0.001
Comments (0)