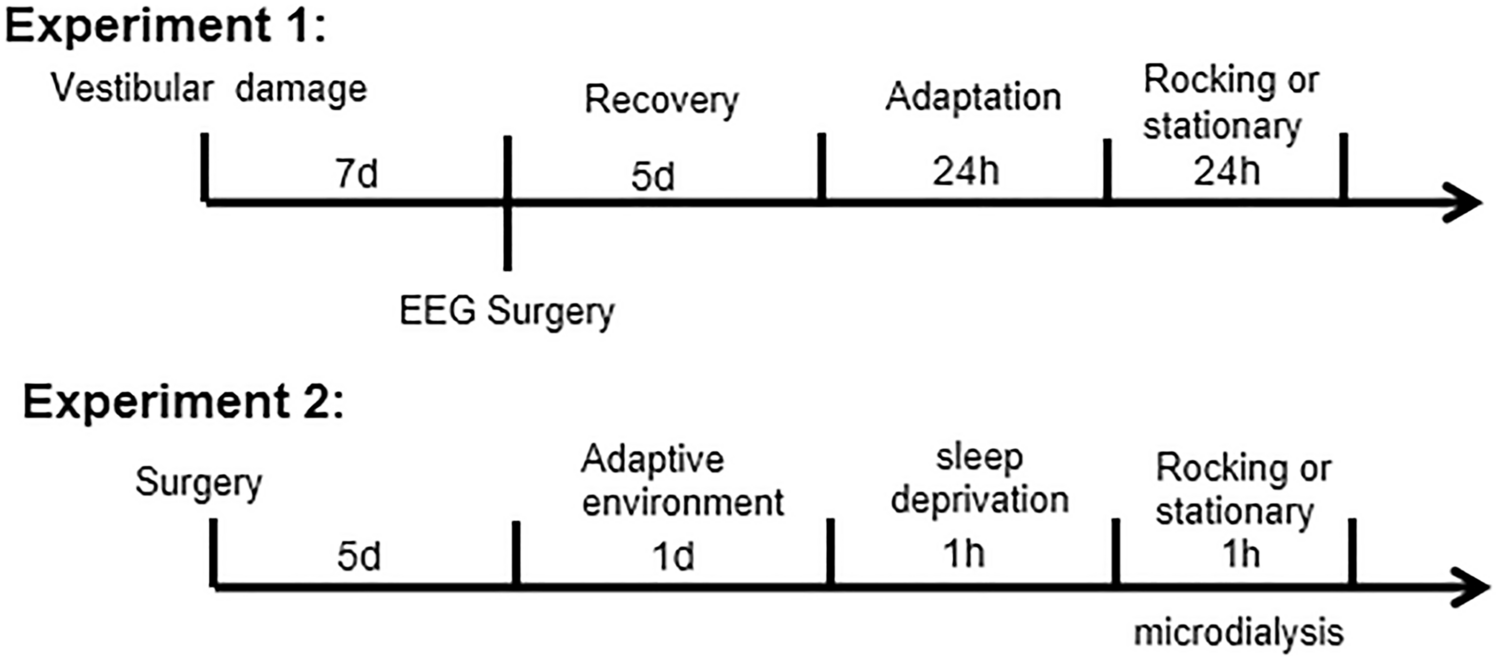
The polysomnography (PSG) system can visually determine the sleep and wakefulness status of rats by EEG. In this experiment, after the rats were given 1.5 HZ uniform rocking motion, the normal rocking group had more NREM sleep and less wakefulness than the vestibular disrupted rocking group, which indicated that the sleep-promoting effect caused by uniform rocking motion was felt by the peripheral vestibular receptors and transmitted by the vestibular nerve pathway, and there was a clear neural pathway between the vestibular and sleep-promoting centers. However, balance is the result of the interplay of peripheral vestibular, visual, and proprioceptive senses, and it cannot be ruled out that visual and proprioceptive senses can respond to swaying, and the subsequent role of visual and proprioceptive senses on the vestibule needs to be considered independently. This is clearly shown as the ratio of REM sleep to total sleep (TS) [11]. We have not replicated the observation of 1.5 Hz sway in mice, probably due to different mechanical characteristics of the vestibular organs. The otolithic membrane can generate shear forces under gravity or inertia that cause cilia to oscillate, following which the cilia of the vestibular hair cells are deflected by tugging, the cellular ion channels open or close, and then signals are transmitted synaptically to the vestibular nerves, which are detected by the brain to perceive movement. Although mice and rats are the same species, they differ in the area of the otolithic membrane, the number of hair cells, and the size of the hair cells [18]. Acceleration is the same, but the otolith organs do not receive the same inertial forces, and the brain receives neural stimuli differently, so the optimal rocking frequencies of mice and rats are likely to differ. Although the results for mice were not replicated in rats, the results for sleep promotion still represent that 178 cm/s2 is still in the optimal range for sleep promotion.
The microdialysis results showed that the content of GABA in the VLPO area began to increase 10 min after the start of rocking movement, peaked after 20 min and began to decrease, and reached the basal value at 40 min. However, there was a slight difference with the time of entering sleep in EEG rats. Rocking to promote sleep may be associated with a transient increase in VLPO, but no significant change in Glu occurred, suggesting that rocking may not be associated with glutamatergic neurons within VLPO.
The increase in GABA content in the VLPO area during uniform rocking movements may be related to the inhibition of its postganglionic neuronal activity, and the EEG wave asynchrony may be related to the possible existence of other neural mechanisms in the ventral lateral preoptic area. Recently, it has been demonstrated that firing of GABAergic neurons in the LMVN (medial vestibular nucleus) underlies stable wakefulness and smooth transition from non-REM sleep to fast sleep, and viral tracing was used to show a direct link between the GABAergic vestibular nucleus and VLPO. This explains rocking can affect GABAergic neurons [19]. Other potential pathways may also indirectly affect sleep. For example, the sleep-promoting mechanisms of AD are in the dorsal forebrain (BF), cerebral cortex and hippocampus, and extracellular levels of AD increase with awake and down time during sleep extending [20]. AD levels are regulated by AD kinase in astrocytes [18, 21]. Injection of AD receptor agonists into the brain increases NREM sleep, while AD antagonists (e.g., caffeine) promote awakening [22]. Rocking may cause sleep by increasing the accumulation of AD in the sleep nucleus. Bilateral vestibular damage in rats can significantly induce theta wave power [23], and patients with bilateral vestibular damage were significantly anxious, but significant reduction in anxiety levels after moderate galvanic vestibular stimulation treatment in healthy young adults [24]. This confirms the modulatory effect of vestibular stimulation on mood state, emotional control, and anxiety levels. This effect of anxiolytic levels may indirectly promote an increase in sleep with a decrease in sleep latency time.
Glu and GABA are among the important neurotransmitters in the central nervous system that regulate sleep–wake, and are important in the normal physiological state of the body [25]. Glu is a widely distributed excitatory neurotransmitter in the brain, which has an important role in maintaining the excitatory state of the cerebral cortex, and we expect its alteration in the constant rocking movement state to be different corresponding to alteration of inhibitory neurotransmitter GABA. The excitatory neurotransmitter Glu content in this experiment did not show significant changes in normal awake rats during peripheral vestibular stimulation and in the group without peripheral vestibular stimulation, so the specific mechanism of whether Glu delivers nerve impulses to vestibular receptors in the vestibular nerve conduction pathway and exerts excitatory effects in the neuromodulatory axis needs to be further investigated. It may be related to the low distribution of Glu neurons in VLPO, but it cannot be excluded that in other nuclei, Glu improves sleep by swaying, and sleep is produced by the joint action of multiple neurotransmitters in multiple nuclei, disrupting neurons in individual parts of the ascending wakefulness system, including cholinergic neurons in the basal forebrain, or multiple neurons such as (basal forebrain cholinergic neurons, orexins neurons and locus coeruleus neurons) also do not have a significant effect on the amount of waking sleep [26, 27]. Therefore, it needs to be further investigated in relation to several neural clusters, such as VLPO projecting to locus coeruleus, histaminergic neurons in the posterior hypothalamic nodal papillary nucleus (TMN), cholinergic neurons in the lateral dorsal tegmental nucleus of the pons, and 5-hydroxytryptaminergic neurons in the dorsal nucleus of the middle suture. These projected areas are inextricably linked to sleep [6]. VLPO active nerves are mainly GABAergic neurons, while GABA neurons of VLPO have subtypes that produce Galanin [28] which is also closely related to sleep [29]. The specific GABAergic neurons in VLPO are able to promote NREM sleep and reduce wakefulness [30], which is similar to our results.
Insomnia can cause decreased quality of life, fatigue, bad moods, decreased learning and memory functions, decreased immune function, and even lead to various diseases. For people with insomnia, there is no proven method other than taking benzodiazepine anti-insomnia medications. Although there have been many studies on sleep stimulation, there are few data to support the use of non-pharmacological treatments for insomnia. Our study shows that moderate amounts of homogeneous rocking movement stimulation can cause an increase in the content of the inhibitory neurotransmitter GABA in the VLPO region, effectively promoting sleep, and the results can be expected to bring value to the combined clinical application.



Comments (0)