
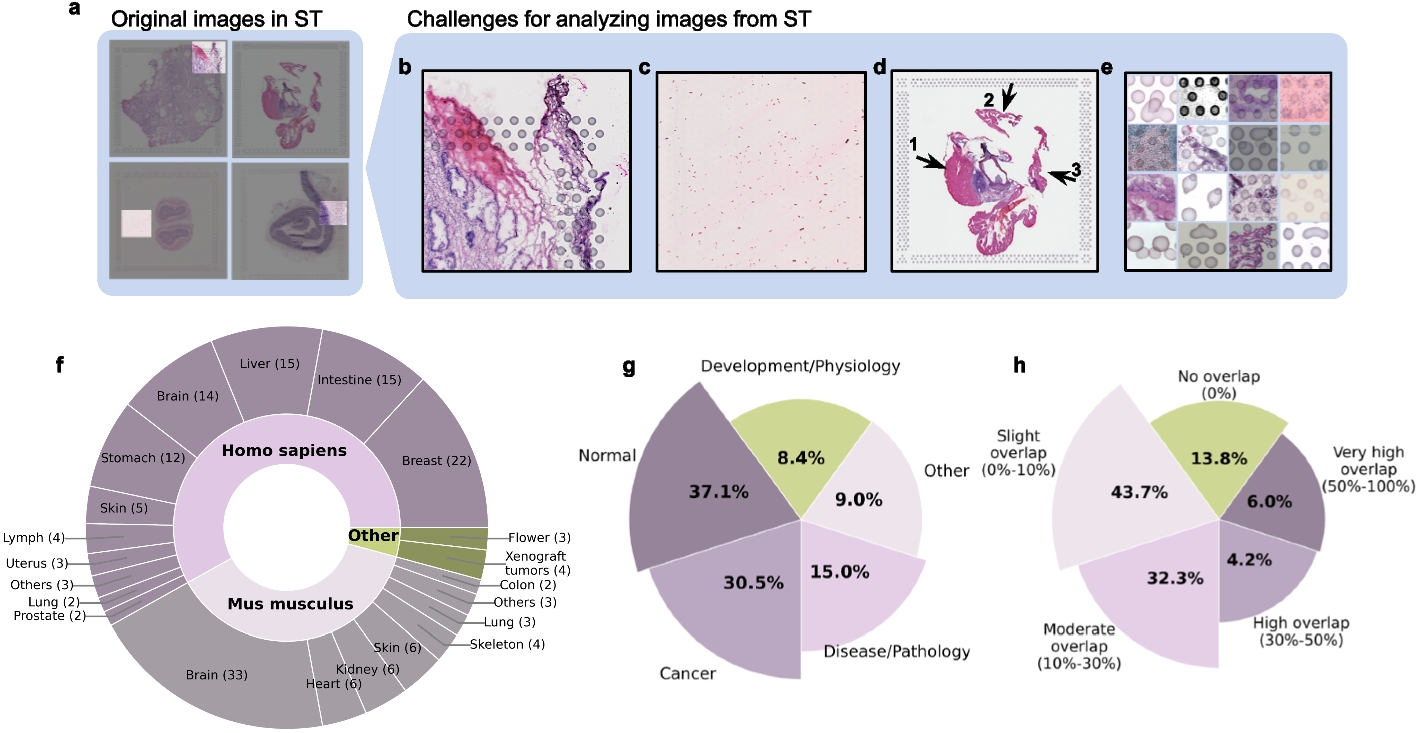
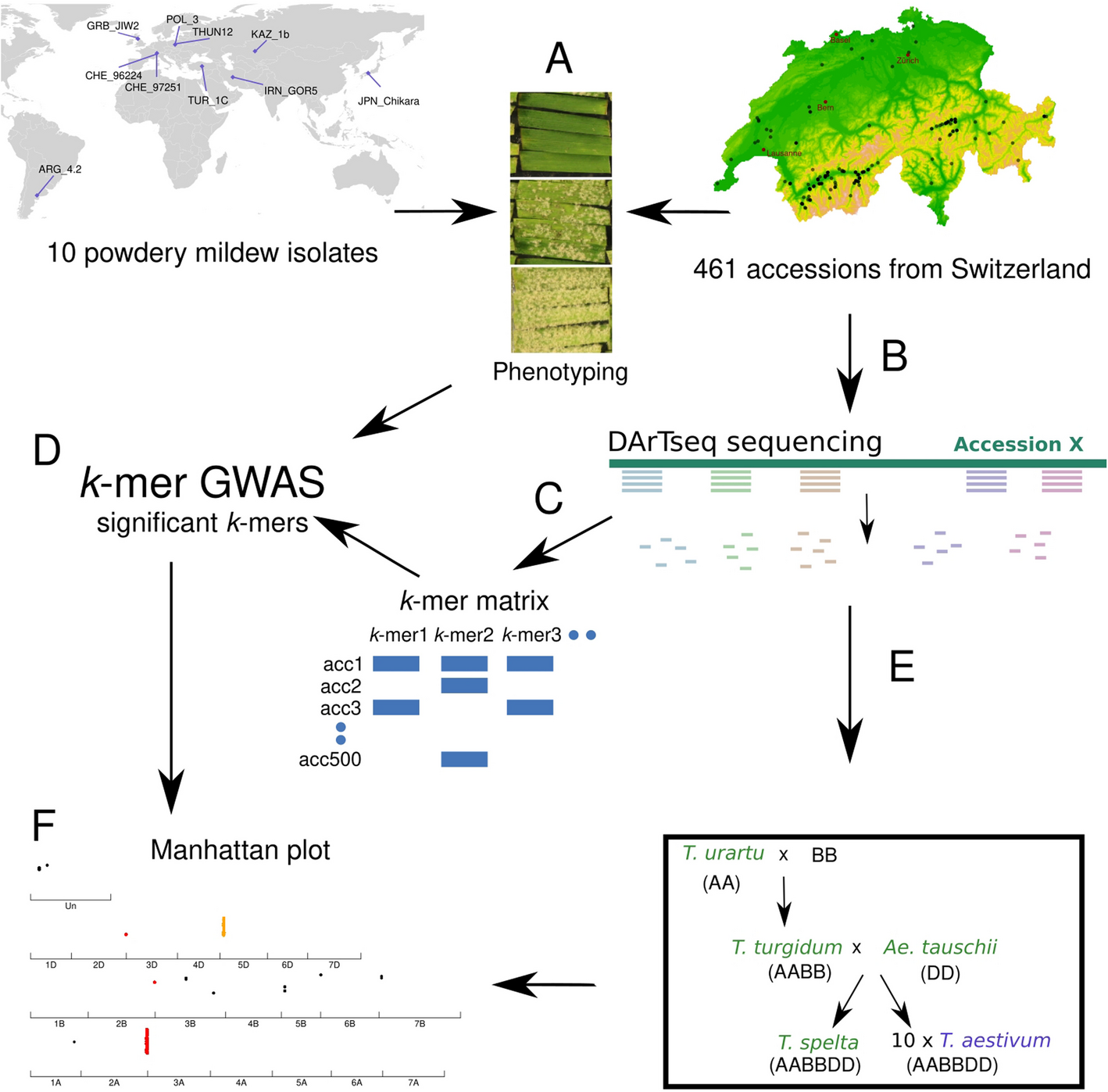
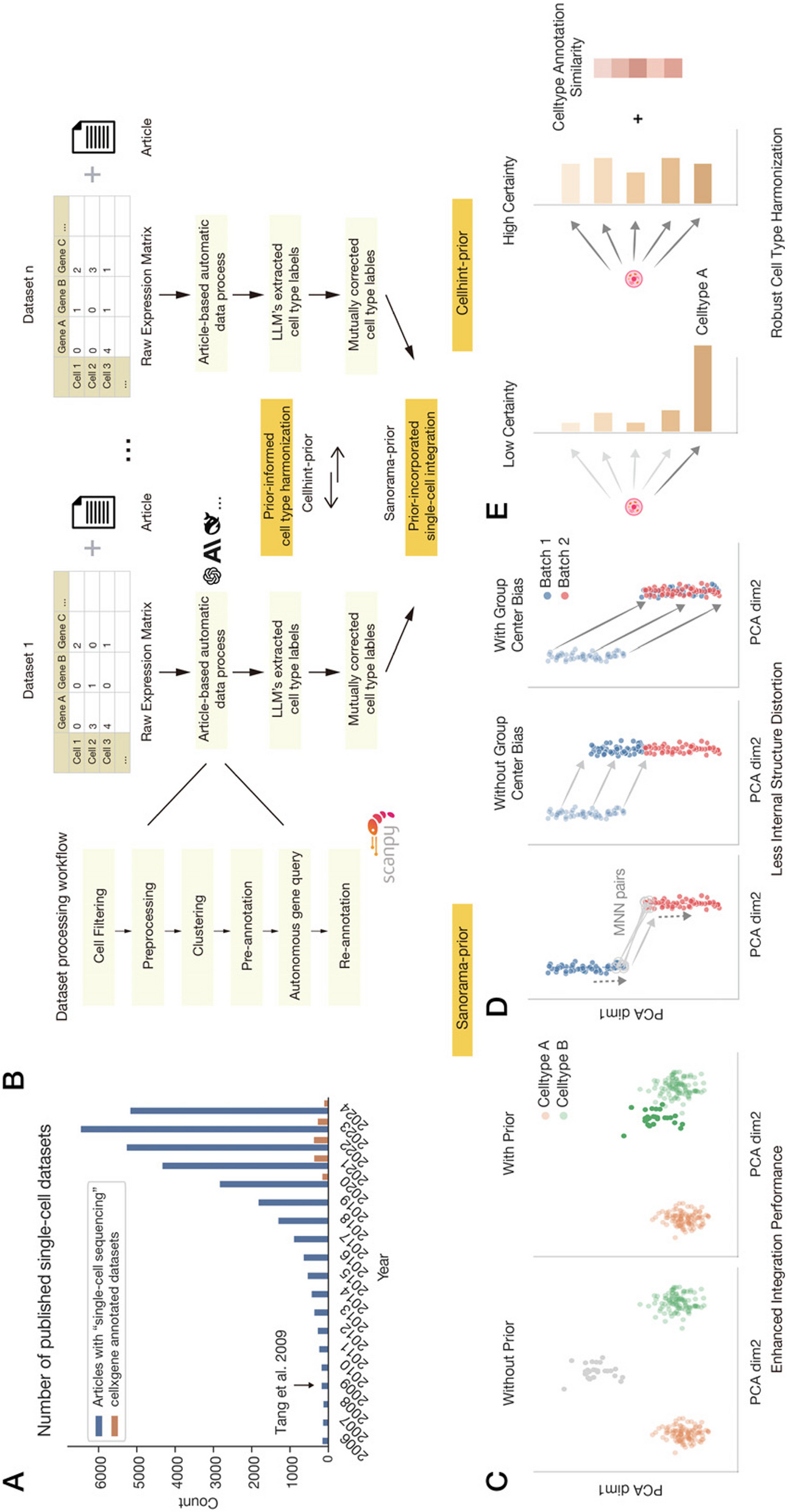
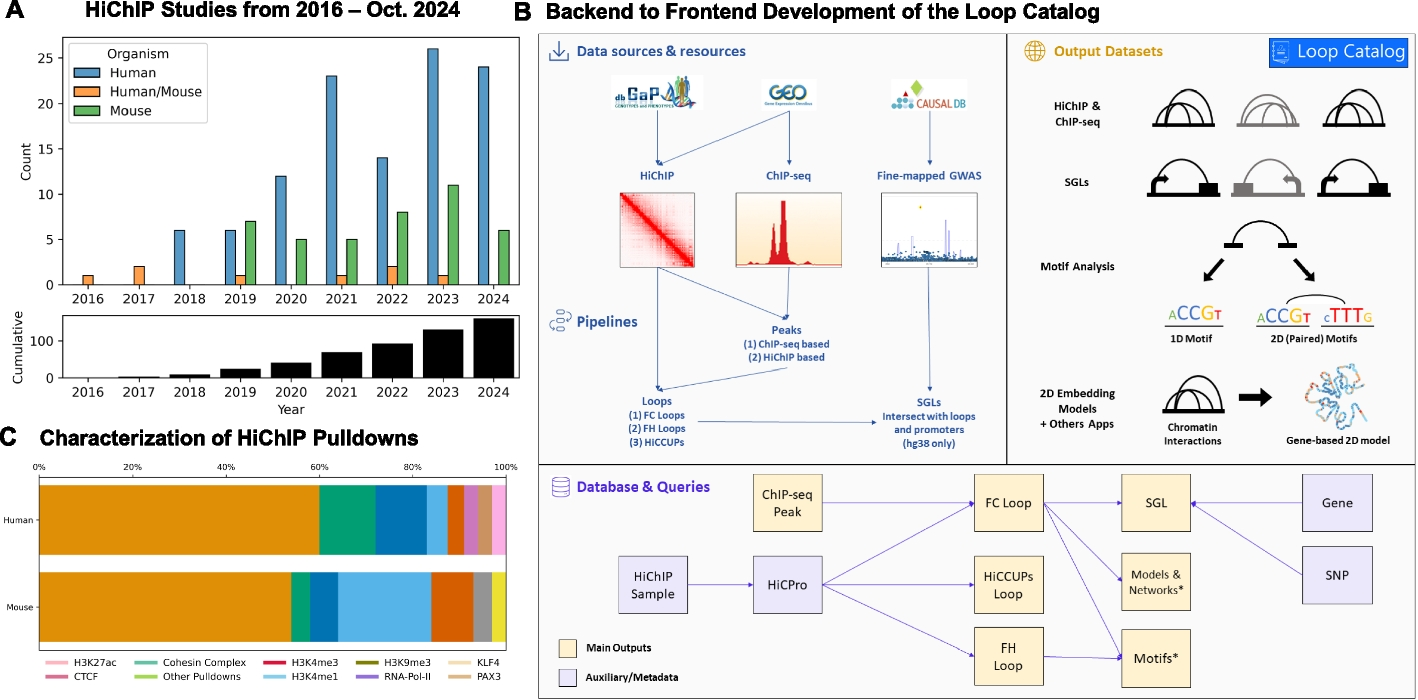
Remember me

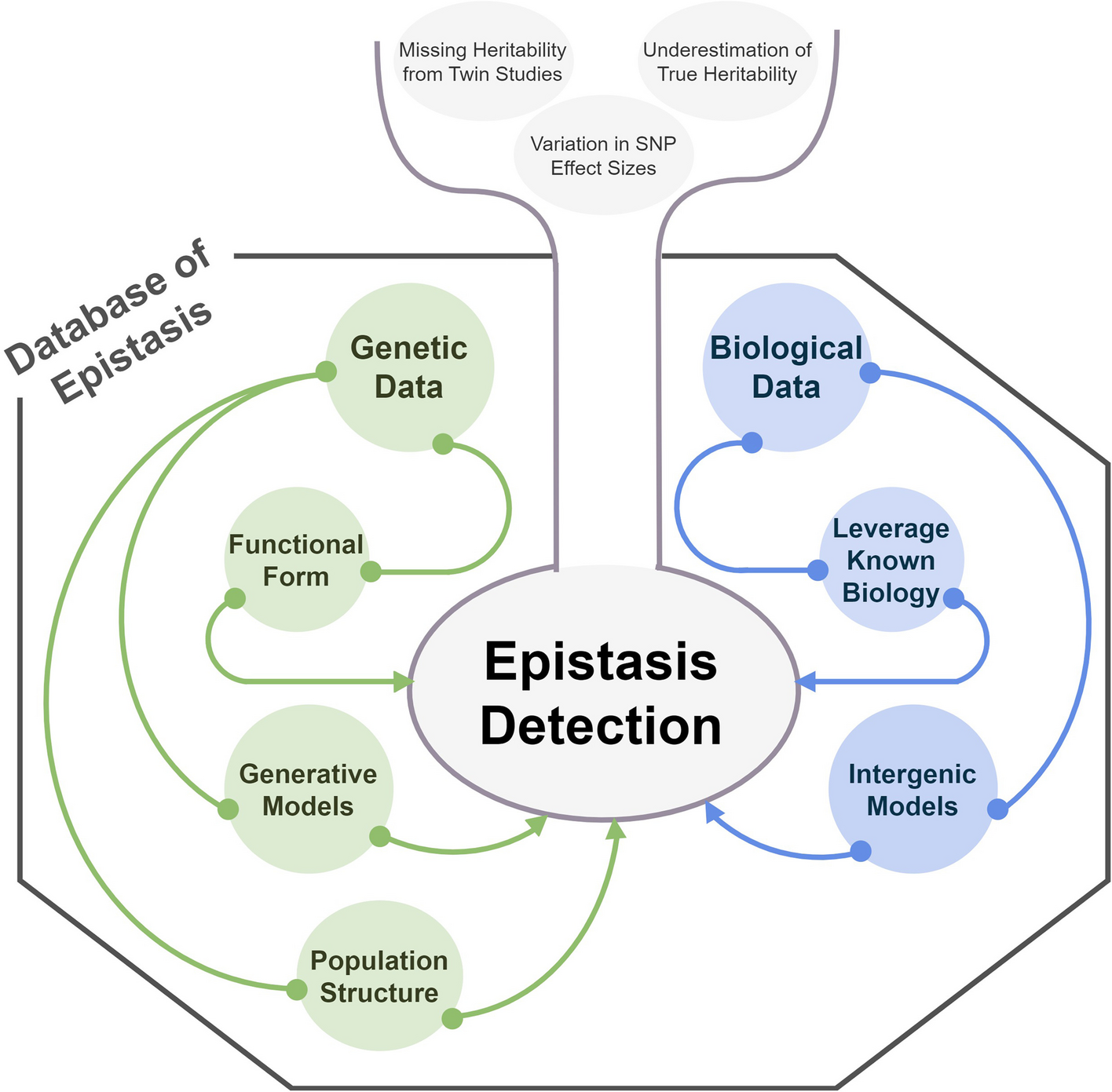

To investigate the metabolic regulatory network in response to UV-B stress in rice seedlings, we collected leaf samples from 160 rice varieties grown under normal and high UV-B conditions, respectively (Additional file 2: Table S1), and analyzed them using an ultrahigh-performance liquid chromatography-tandem mass spectroscopy (UPLC-MS/MS)-based widely targeted metabolic profiling method as described previously [35, 36]. A total of 810 metabolic features were determined; among them, 143 were confirmed by comparison with standards and 326 were putatively annotated, including alkaloids, amino acids, fatty acids, flavonoids, hormones, nucleic acids, others, phenolamines, polyphenols, terpenes, and vitamins (Additional file 1: Fig. S1; Additional file 2: Table S2). Principal component analysis (PCA) of these metabolites showed a clear separation between the normal and UV-B treatment conditions along the first component, while the second component separated the indica and japonica subspecies, consistent with a previous report [28]. Interestingly, this subspecies separation was more pronounced under UV-B conditions, indicating a possible role and mechanism of metabolite regulation in subspecies differentiation and the stress response (Fig. 1a). Furthermore, differentially accumulated metabolite analysis between treated and control samples indicated that most (436/810) of the metabolites were significantly influenced by UV-B stress, namely 47.5% (385/810) metabolites showed upregulated patterns and 6.3% (51/810) displayed downregulated patterns (|Fold Change|≥ 5, P < 0.05) under the UV-B stress compared with normal conditions (Fig. 1b). Notably, these annotated and upregulated metabolites mainly belonged to amino acids, fatty acids, and flavonoids (Fig. 1c; Additional file 1: Fig. S2), indicating these metabolites might confer and control rice UV-B response and tolerance. Additionally, the cluster dendrogram based on differential accumulation of the metabolites showed two distinct groups under the UV-B conditions, in line with the PCA results (Fig. 1d). Collectively, these results suggest that the identified UV-B-responsive metabolites could efficiently reflect and even play substantial roles in the response and tolerance to UV-B in rice.
Fig. 1
Metabolic differential accumulation of rice seedling leaves under normal and UV-B conditions. a Principal component analysis (PCA) of metabolites of rice seedling leaves under normal and UV-B conditions. b Volcano plot displaying upregulated and downregulated metabolites under UV-B conditions compared to those under the normal conditions; the red dot represents a metabolite with a |Fold Change|≥ 5 and the blue dot represents a metabolite with a |Fold Change|≤ 0.2, P ≤ 0.05. c Percentage of differential metabolites identified under UV-B conditions compared to those under the normal conditions. d Neighbor-joining tree of 160 rice varieties based on 810 metabolites under normal and UV-B conditions
UV-B-responsive transcriptomic profiling in riceTo better understand the mechanism underlying the UV-B-responsive metabolites in rice, we selected some rice varieties for UV-B tolerance analysis and found that different rice varieties showed distinct difference to UV-B tolerance (Additional file 1: Fig. S3). Further four rice varieties (two conventional varieties, ZH11 and ZS97, medium susceptible and medium resistant to UV-B stress respectively; a UV-B resistant variety, Lemon; and a UV-B susceptible variety, Dular) were used for transcriptome analysis under normal and UV-B conditions. PCA showed greater differences in gene expression levels between the normal and UV-B treatment conditions than among the four rice varieties, with the first component explaining 22.5% of the total variance (Fig. 2a). Comparative analysis of differentially expressed genes (DEGs) in these rice varieties under UV-B treatment revealed 9029 (3995 up-regulated and 5034 down-regulated), 7596 (3047 up-regulated and 4549 down-regulated), 7870 (3831 up-regulated and 4039 down-regulated), and 9296 (5251 up-regulated and 4045 down-regulated) DEGs in ZH11 (Additional file 2: Table S3), ZS97 (Additional file 2: Table S4), Dular (Additional file 2: Table S5), and Lemon varieties (Additional file 2: Table S6), respectively, compared with the corresponding expression levels under the normal conditions (Fig. 2b). Among these DEGs, 1296 and 992 genes were commonly up-regulated and down-regulated by UV-B, respectively (Fig. 2c).
Fig. 2
Global transcriptome responses in seedling leaves of four rice varieties under normal and UV-B conditions. a PCA of transcriptome reprogramming in seedling leaves of four rice varieties namely ZH11, ZS97, Lemon, and Dular under normal and UV-B conditions. b Number of differentially expressed genes (DEGs) in seedling leaves of four rice varieties under UV-B conditions; red, up-regulated genes and blue, down-regulated genes. c Venn diagrams showing overlap of DEGs in seedling leaves of four rice varieties under UV-B conditions. d KEGG pathway enrichment analysis for DEGs in seedling leaves of four rice varieties under UV-B conditions
To gain insight into biological processes involved in the response to UV-B stress, Kyoto Encyclopedia of Genes and Genome (KEGG) pathway enrichment analysis was performed on the 2288 UV-B-responsive DEGs in these four rice varieties. The set of 1296 commonly up-regulated DEGs was mainly involved in phenylalanine, tyrosine, and tryptophan biosynthesis, phenylpropanoid metabolism, and flavonoid biosynthesis (Fig. 2d). The 992 DEGs uniquely down-regulated under UV-B stress were mostly enriched in glycine, serine, and threonine metabolism, glyoxylate and dicarboxylate metabolism, and cysteine and methionine metabolism (Fig. 2e). These results imply that the regulation of phenylpropanoid, tyrosine, and tryptophan biosynthesis plays crucial roles in the response to UV-B stress in rice, consistent with the metabolome profiling results under the UV-B conditions.
Genetic basis of differential metabolites in response to UV-B in riceTo investigate the genetic basis of the differential metabolites identified in response to UV-B stress in rice seedlings, we calculated the coefficient of variation of 810 metabolites in the 160 rice varieties under normal and UV-B conditions. The results showed a higher proportion of variance in rice varieties under the UV-B conditions, implying that the metabolites responding under UV-B treatment exhibited significant genetic diversity (Additional file 1: Fig. S4). Subsequently, mGWAS was performed in the normal and UV-B conditions to identify significant loci associated with the differential metabolites (Fig. 3a, b; Additional file 1: Fig. S5). A total of 3127 and 4149 significantly induced loci were detected under normal and UV-B conditions, respectively, indicating their relevance for screening candidate genes to explore the genetic basis of differential metabolites in response to UV-B stress.
Fig. 3
Overall mGWAS results of the metabolic features in rice seedling leaves under normal and UV-B conditions. a Genomic distribution of mGWAS results for the metabolic features in rice seedling leaves under normal and UV-B conditions; all metabolite–SNP associations with P ≤ 5.0 × 10−6 are plotted; metabolites were grouped into seven categories and marked with different colors namely a-e. b Venn diagram showing the number of significant loci in mGWAS result under normal and UV-B conditions; P ≤ 5.0 × 10−6
To explore the functions of genes associated with UV-B stress in these significant loci, Gene Ontology (GO) enrichment and KEGG pathway enrichment analyses were conducted. The majority of the candidate genes were involved in biological processes, cellular components, and molecular functions by GO analysis (Additional file 1: Fig. S6; Additional file 2: Table S7). Additionally, KEGG pathway analysis revealed that the genes that were induced under normal conditions were primarily involved in amino acid metabolism, fatty acid metabolism, and glycolysis, whereas those induced under the UV-B conditions were predominantly involved in phenylpropanoid biosynthesis, phenylalanine metabolism, and starch and sucrose metabolism (Additional file 1: Fig. S7; Additional file 2: Table S8). These analyses supported that the regulation of phenylpropanoid biosynthesis and phenylalanine metabolism likely play critical roles in the UV-B response and tolerance in rice. Combined with the results of the transcriptome data, candidate genes involved in amino acid and flavonoid metabolism were further determined and identified.
OsMYB44 regulates tryptamine accumulation contributing to UV-B tolerance in riceN-cinnamoyl tryptamine, a derivative of tryptamine, exhibited distinct loci under the UV-B stress conditions, as demonstrated by association analysis (Fig. 4a; Additional file 2: Table S9). Candidate gene selection revealed that locus L1, SNP 900647643 (P = 9.8 × 10−9), was situated 7 kb upstream of OsMYB44, encoding a transcription factor in the MYB family (Fig. 4b; Additional file 1: Fig. S8). Moreover, induction analysis showed that UV-B significantly induced the expression of OsMYB44 in rice, along with genes associated with tryptamine biosynthesis, namely OsTSα, OsTSβ, OsTDC1, OsTDC3, OsTHT1, and OsTBT2 (Additional file 1: Fig. S9).
Fig. 4
Identification and functional determination of OsMYB44 in rice. a Manhattan plots showing the GWAS result on cinnamoyl tryptamine content in the leaves of 160 rice varieties in normal and UV-B conditions. b Association between polymorphic sites within the OsMYB44 loci and cinnamoyl tryptamine content (each dot represents a polymorphic site); representation of pairwise r2 values (a measure of LD) among polymorphic sites in OsMYB44. c, d Phenotypes of 4-week-old OsMYB44-overexpressing (c) and OsMYB44-CRISPR (d) rice seedling lines and wild-type seedling lines before UV-B treatment and after 72 h UV-B treatment for 10-day recovery. e Survival rate of 4-week-old OsMYB44 transgenic rice seedlings and wild-type seedlings after UV-B treatment for 10-day recovery. f Shoot fresh weight of OsMYB44 transgenic rice seedlings and wild-type seedlings after UV-B treatment for 10-day recovery. g Subcellular localization of OsMYB44 in tobacco leaves; OsGhd7-RFP used as a nuclear marker, bars 20 μm. h Yeast-one hybrid assays showing the OsMYB44 bound to OsTSα and OsTSβ promoter, an empty vector expressing the AD domain as the negative control. i Bar graphs showing the activity of the ProOsTSα:LUC reporter, the ProOsTSβ:LUC reporter, and the ProOsTDC1:LUC reporter in tobacco leaves. LUC activity value was normalized to REN activity as an internal control; REN, renilla luciferase; LUC, firefly luciferase. j Electrophoretic mobility shift assay (EMSA) analysis of OsMYB44 binding to the MBSI motif in the OsTSβ promoters. Fifty-fold molar excesses of unlabelled probes were used in the competition assay. k Relative expression levels of OsTSβ in OsMYB44 transgenic rice seedling leaves and wild-type seedling leaves under normal and UV-B conditions by qRT-PCR. l Relative content of tryptamine and tryptamine derivatives in OsMYB44 transgenic rice seedling lines under UV-B conditions. Data in e, f, i, and k are shown as the means ± SD (n = 3) and *, P < 0.05 and **, P < 0.01 (Student’s t-test) indicate significant differences
To confirm the function of OsMYB44 in rice, we first analyzed its potential role in UV-B stress tolerance in OsMYB44-overexpressing and CRISPR-mediated OsMYB44-silenced rice seedling lines. The overexpression lines exhibited UV-B stress tolerance as evidenced by their green leaves under UV-B exposure (Fig. 4c, e). In contrast, the OsMYB44-CRISPR rice seedling lines showed clear withering symptoms and lower survival rates compared to those of wild-type seedlings after UV-B treatment (Fig. 4d, e). Consistent with these findings, the shoot fresh weight of the OsMYB44-overexpressing lines was higher than that of the wild type under UV-B exposure (Fig. 4f), indicating that OsMYB44 positively regulates UV-B tolerance in rice.
Further examination of the mechanism underlying OsMYB44-mediated UV-B tolerance in rice revealed that OsMYB44 as a functional transcription factor, directly binding to the promoters of OsTSα and OsTSβ (Fig. 4g, h) and significantly activating ProOsTSα:LUC, ProOsTSβ:LUC, and ProOsTDC1:LUC reporter activities (Fig. 4i). The electrophoretic mobility shift assay results further confirmed that OsMYB44 directly binds to the MBSI element in the OsTSβ promoter (Fig. 4j). Moreover, the expression levels of OsTSα, OsTSβ, OsTDC1, and OsTHT1 were significantly increased in OsMYB44-overexpressing seedling lines, particularly after UV-B treatment (Fig. 4k; Additional file 1: Fig. S10). Consequently, the contents of N-cinnamoyl tryptamine, N-benzoyl tryptamine, N-p-coumaroyl tryptamine, and tryptamine significantly increased in these lines (Fig. 4l). These results suggest that OsMYB44 regulates tryptamine biosynthesis to enhance UV-B tolerance in rice. To further investigate the response of tryptamine to UV-B stress in rice, the rice seedlings were treated with exogenous tryptamine, resulting in a significant enhancement of their UV-B stress tolerance compared to that in the control conditions (Additional file 1: Fig. S11a–c). The survival rate of rice seedlings treated with tryptamine was considerably higher than that in the control conditions, especially after 48 h of UV-B exposure (Additional file 1: Fig. S11d). Therefore, we can conclude that OsMYB44 promotes UV-B tolerance in rice by regulating tryptamine accumulation.
OsUVR8 interacts with OsMYB110 to regulate flavonoid accumulation contributing to UV-B tolerance in riceC-hexosyl-luteolin O-hexoside, a flavonoid derivative, displayed distinct genetic loci associated with UV-B stress, as revealed in the association analysis (Fig. 5a). Candidate gene selection identified locus L2, SNP421222223 (P = 1.9 × 10−10), on chromosome 4 positioned upstream of OsUVR8, which is a regulator encoding a protein with multiple RCC1 domains involved in chromosome condensation (Fig. 5b; Additional file 2: Table S10). Phylogenetic analysis demonstrated the similarity of UVR8 amino acid sequences between monocotyledons and dicotyledons (Fig. 5c). Co-expression analysis revealed significant enrichment of co-expressed genes with OsUVR8 in flavonoid biosynthesis (Fig. 5d). Furthermore, the overexpression of OsUVR8 in rice seedling lines led to the upregulation of OsPAL1, Os4CL5, and OsCHS expression, while the downregulated expression of these genes was observed in OsUVR8-CRISPR rice seedling lines (Additional file 1: Fig. S12a–e). Accordingly, the content of several flavonoids such as methyl apigenin C-hexoside, C-hexoside-apigenin O-p-coumaroyl hexoside, chrysoeriol 7-O-rutinoside, and C-hexoside-chrysoeriol O-hexoside significantly increased in OsUVR8-overexpressing rice seedling lines (Additional file 1: Fig. S12f). These findings indicate that OsUVR8 plays a regulatory role in flavonoid accumulation in rice. Additionally, the OsUVR8-overexpressing rice seedling lines exhibited enhanced tolerance to UV-B stress, with notably higher survival rates, while OsUVR8-CRISPR rice seedling lines displayed increased sensitivity to UV-B stress, with lower survival rates compared to those of wild-type rice lines after UV-B treatment (Fig. 5e, f). Notably, the shoot fresh weight was higher in OsUVR8-overexpressing rice seedling lines compared to that of wild-type rice lines following UV-B treatment (Fig. 5g). These results indicated that OsUVR8 is involved in flavonoid accumulation and UV-B tolerance in rice.
Fig. 5
Identification and functional determination of OsUVR8 and OsMYB110 in rice. a Manhattan plots showing the GWAS result on C-hexosyl-luteolin O-hexoside content in the leaves of 160 rice varieties under normal and UV-B conditions. b Association between polymorphic sites within the OsUVR8 loci and C-hexosyl-luteolin O-hexoside content (each dot represents a polymorphic site); representation of pairwise r2 values (a measure of LD) among polymorphic sites in OsUVR8 containing many RCC1 domain. c Phylogenetic comparison of OsUVR8 in plants; bootstrap values from 1000 resamplings are indicated. The bar represents 0.5 amino acid substitutions per site. d Co-expressed analysis of OsUVR8 in rice. e Phenotypes of 4-week-old OsUVR8 transgenic rice seedling lines and wild-type seedling lines before UV-B treatment and after 72 h UV-B treatment for 10-day recovery. f Survival rate of 4-week-old OsUVR8 transgenic rice seedlings and wild-type seedlings after UV-B treatment for 10-day recovery. g Shoot fresh weight of OsUVR8 transgenic rice seedlings and wild-type seedlings after UV-B treatment for 10-day recovery. h Yeast two-hybrid assay for interaction between OsUVR8 and OsMYB110, empty vectors pGADT7 as a control in UV-B conditions. i BiFC analysis for interaction between OsUVR8 and OsMYB110 in tobacco, full-length OsUVR8 fused to C-terminal YFP (OsUVR8-cYFP) and full-length OsMYB110 fused to N-terminal YFP (nYFP-OsMYB110); OsUVR8-cYFP and nYFP as controls, bars 20 μm. j In vitro pull-down assays showing OsUVR8 interacting with OsMYB110. His-OsUVR8 were incubated with GST-MYB110 and pulled down from GST-MYB110 conjugated GST beads. The eluates were analyzed by immunoblots with anti-His and anti-GST antibodies respectively. k Bar graphs showing the activity of ProOsCHS:LUC reporter in tobacco leaves. LUC activity value was normalized to REN activity as an internal control; REN, renilla luciferase; LUC, firefly luciferase. Data in f, g, and k are shown as the means ± SD (n = 3) and *, P < 0.05 and **, P < 0.01 (Student’s t-test) indicate significant differences
To investigate the role of OsUVR8 in flavonoid accumulation and UV-B tolerance in rice, a yeast two-hybrid screen was conducted with and without UV-B treatment to identify proteins interacting with OsUVR8. OsMYB110 was identified as an interacting partner in the presence of UV-B treatment (Fig. 5h). Bimolecular fluorescence complementation and GST pull-down assays further confirmed the interaction between OsUVR8 and OsMYB110 in vivo and in vitro (Fig. 5i, j). Moreover, transient transcription analysis demonstrated that both OsUVR8 and OsMYB110 individually activated the activity of ProOsCHS:LUC, with a more pronounced effect observed when OsUVR8 was co-expressed with OsMYB110, particularly under the UV-B conditions (Fig. 5k).
To further underline the involvement of OsMYB110 in flavonoid accumulation and UV-B tolerance in rice, the UV-B-induced expression patterns of OsMYB110 and key genes involved in flavonoid biosynthesis, including OsPAL1, Os4CL5, OsCHS, OsF3H, OsFLS, and several OsUGTs, were analyzed, revealing significant upregulation under UV-B stress (Additional file 1: Fig. S13). Yeast one-hybrid and transient transcription assays demonstrated the binding of OsMYB110 to the promoters of OsPAL1, Os4CL5, and OsCHS, along with remarkable activation of ProOsPAL1:LUC, ProOs4CL5:LUC, ProOsC4H:LUC, and ProOsCHS:LUC reporter activities (Additional file 1: Fig. S14a, b). Moreover, OsPAL1, Os4CL5, OsCHS, OsF3H, and OsCGT showed significant upregulation in OsMYB110-overexpressing rice seedling lines, particularly after UV-B treatment (Additional file 1: Fig. S14c). Consequently, higher levels of most flavonoids were observed in OsMYB110-overexpressing rice seedling lines under both normal and UV-B conditions (Additional file 1: Fig. S14d). Additionally, OsMYB110-overexpressing rice seedling lines exhibited reduced UV-B-induced damage and higher survival rates compared to those of the wild-type rice lines (Additional file 1: Fig. S14e, f). Similarly, the shoot fresh weight of OsMYB110-overexpressing rice seedling lines was significantly higher than that of the wild-type rice lines after UV-B treatment (Additional file 1: Fig. S14g). These findings provide evidence that OsUVR8 interacts with OsMYB110 to regulate flavonoid accumulation and enhance UV-B tolerance in rice.
Natural variation of OsUVR8 contributes to UV-B tolerance in riceTo investigate the relationship between OsUVR8 haplotypes and UV-B intensity in a diverse collection of rice accessions in the world, the nucleotide polymorphisms of OsUVR8 were analyzed in 4275 rice accessions based on information from RiceVarMap V2.0 (http://ricevarmap.ncpgr.cn/). Twenty-five SNPs and insertion-deletions were categorized into two distinct groups, namely group A and group B, which encompassed thirteen haplotypes. Group A consisted of haplotypes H3, H4, H5, H6, H7, and H10, and the remaining haplotypes were assigned to group B (Fig. 6a, b). Interestingly, rice varieties with haplotypes in group A (referred to as OsUVR8groupA) exhibited significantly high levels of most flavonoids (61 upregulared patterns in 70 differentially accumulated flavonoids) with P < 0.05, whereas rice varieties with haplotypes in group B (referred to as OsUVR8groupB) displayed notably low levels of most flavonoids (Fig. 6c). Transient assays further revealed that the promoter activity of OsUVR8groupA was substantially higher than that of OsUVR8groupB in tobacco (Fig. 6d). Moreover, geographical distribution analysis demonstrated that approximately 6% of rice varieties with OsUVR8groupB and only 1% of varieties with OsUVR8groupB inhabit areas characterized by low and high UV-B intensity (Fig. 6e). These findings suggest that rice varieties with OsUVR8groupA are likely to have been artificially selected for their ability to withstand high UV-B intensity conditions.
Fig. 6
Natural variation and geographical distribution of OsUVR8 in rice varieties. a Haplotype analysis of OsUVR8 in 4275 rice varieties; thirteen haplotypes were detected, and the numbers of rice accessions with thirteen haplotypes were shown. b Phylogenetic tree of the thirteen haplotypes divided into group A (Hap3-Hap7 and Hap10), and group B (Hap1, Hap2, Hap8, Hap9, Hap11, Hap12, and Hap13). The scale bar indicates the average number of substitutions per site for different haplotypes. c Hierarchical clustering showing flavonoid accumulation in rice variety leaves with the two Groups. Red represents high flavonoid content, and blue represents low flavonoid content. d Bar graphs showing the promoter activity of the OsUVR8 with different haplotypes in tobacco. LUC activity value was normalized to REN activity as an internal control; REN, renilla luciferase; LUC, firefly luciferase. e Geographic distribution among rice varieties with group A and group B in this study. Each pie chart size was proportional to the number of rice accessions with different groups in geographic location; red showing high UV-B intensity and blue showing low UV-B intensity
Comments (0)