
Remember me


Gallocyanin is a progressive Nissl stain that has been demonstrated to bind to nucleic acids [2]. It is well established that neurons are known to contain high concentrations of DNA and RNA. The cerebral cortex and subcortical nuclei are distinguished by their high neuron density. The delineation of regional variations in neuron density and arrangement is facilitated by using 400-µm-thick sections (Fig. 1a, b). The telencephalic medullary layer is primarily comprised of oligodendro- and astrocytes, which are virtually devoid of Nissl substance in light microscopic preparations. This results in a faint staining of the telencephalic white matter. However, upon closer examination, the periventricular parts of the central white matter appear lighter, whereas the medullary rays in the gyri appear darker. This phenomenon is particularly evident in the temporal lobe, which is demarcated from the frontal lobe by the lateral sulcus (S l).
Fig. 1
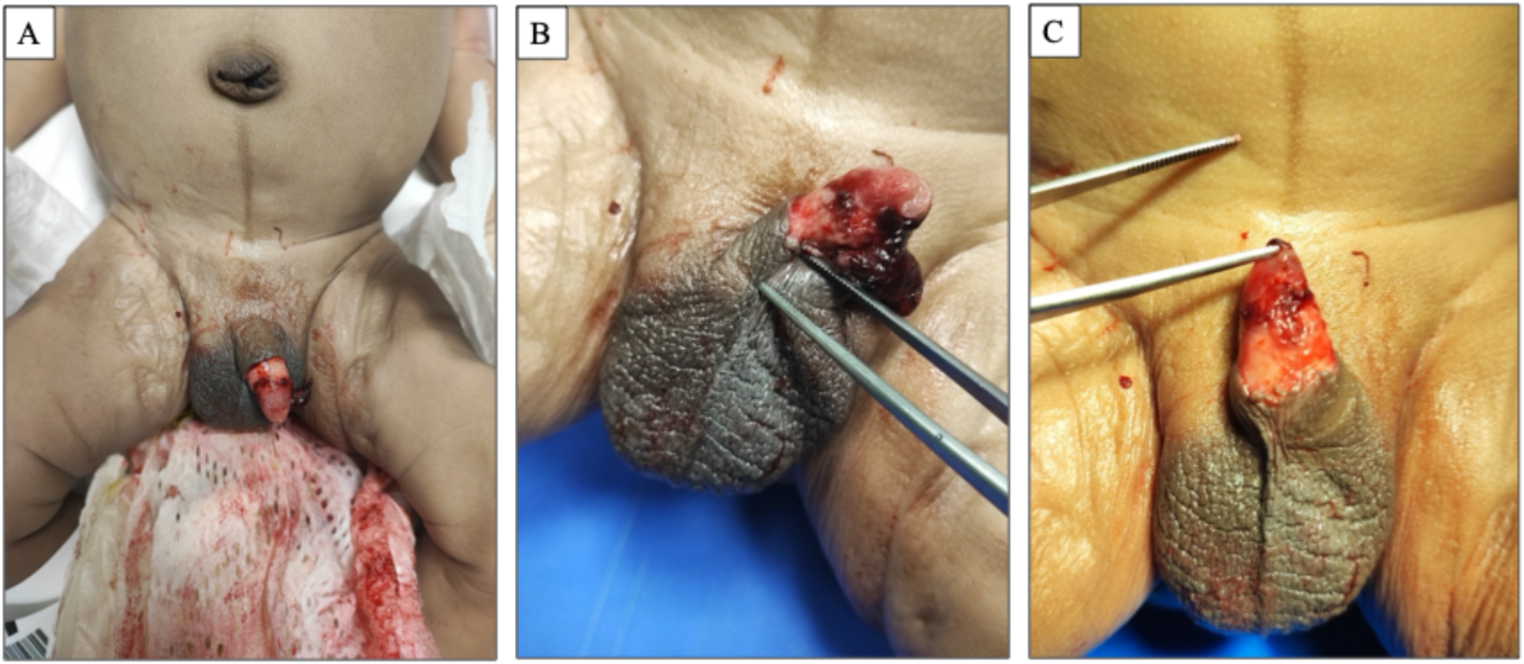
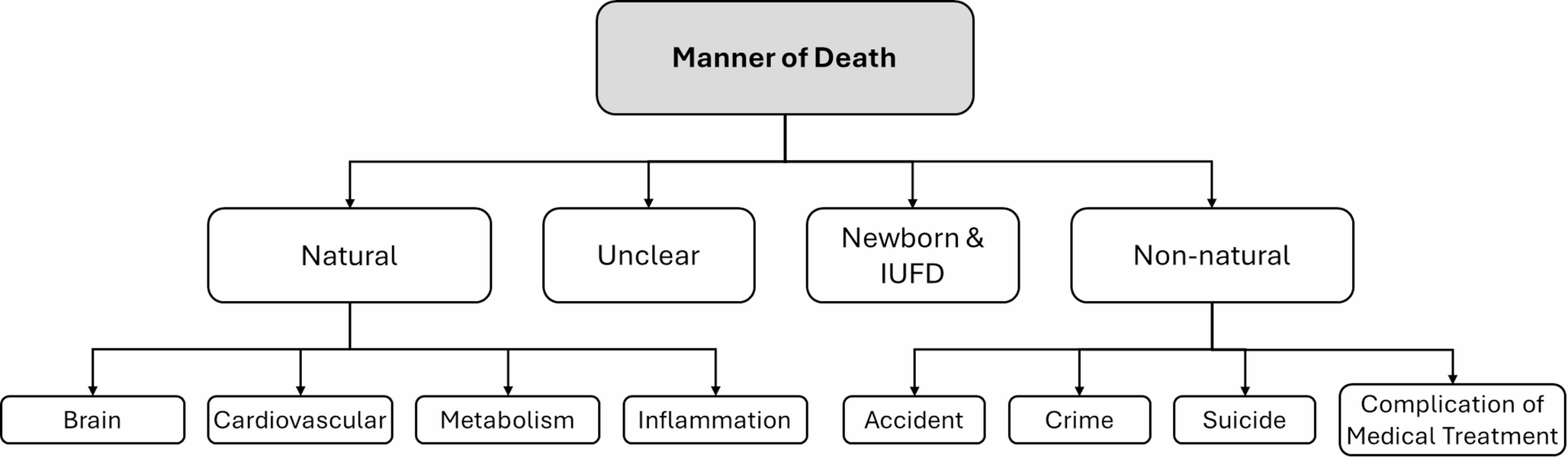
Overview of gallocyanin-stained 400 μm thick frontoparallel brain slices at the level of the mammillary bodies. (a) Case 1 44-year-old man, (b) Case 2 77- year-old man. The asterisks indicate cutting artefacts of unfixed brain tissue
It is imperative to acknowledge that the process of dissecting unfixed brain tissue is susceptible to various forms of tissue deformation, loss, and uneven section thickness. The asterisks in Fig. 1 denote staining differences attributable to artefactual variations in gross section thickness and tissue loss.
Both slices were selected from comparable planes of coronal sections. However, a discernible variation in total size is evident, with the gyri of the 77-year-old male subject exhibiting a reduction in size, accompanied by an expansion of the sulci and the ventricular system (v). A stereologic estimation employing point-counting methods [3] yielded a total hemispheric area of 55.8 cm² vs. 45.5 cm² (-18.6%), 21.2 cm² vs. 17 cm² (-19.9%) of cortical tissue, and 34.6 cm² vs. 28.4 cm² (-17.9%) of subcortical tissue, which encompasses the white matter and subcortical nuclei. The data in brackets quantify the amount of nervous tissue deficit in the 77-year-old male subject compared with the 44-year-old male subject.
The medial temporal lobe comprises corticoid, allo-, periallo-, and isocortical elements. The optic tract (To) courses in close proximity to the amygdaloid complex [4]. The shallow semi-annular sulcus demarcates the boundary between the amygdaloid cortical nucleus (Co) and the anterior hippocampal fields (Fig. 2a).
Fig. 2
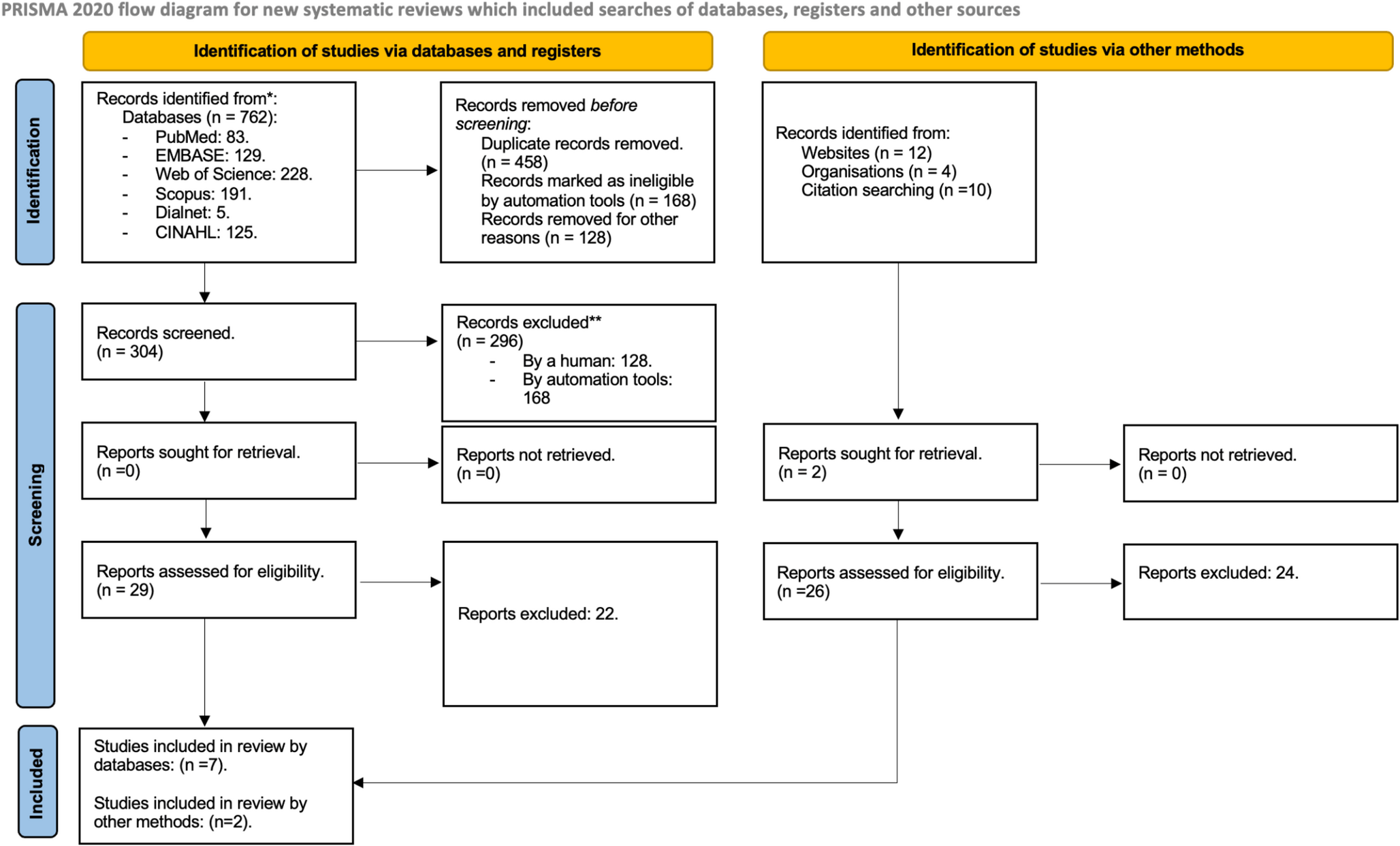
Close-up view of the right mediobasal temporal lobe. (a) Case 1, 44-year-old man, (b) Case 2, 77- year-old man
Individual nuclei of the amygdaloid complex are grouped [5]. The central nucleus (Ce) and the medial nucleus (Me) form the deep centromedial group. They are located in close proximity to the cholinergic and chromophilic nuclear complex of Mesulam’s Ch4p [5], a component of the basal forebrain complex or Meynert’s basal nucleus [6]. In Fig. 2b Ch4i, an additional nuclear group of the basal forebrain complex, can be identified. The superficial cortical amygdaloid region is divided into several sectors. In Fig. 2a, only the cortical nucleus (Co) and its subdivisions could be traced. The basolateral nuclear group consists of the lateral (La), basolateral (BL), basomedial (BM), and paralaminar (PL) amygdaloid nuclei. Nuclear groups and individual amygdaloid nuclei are characterized by specific afferent and efferent connections and functions [7].
The three-layered anterior hippocampal fields (Hip), together with the subicular fields (S) and the entorhinal cortex (R e), constitute the hippocampal formation [8], which extends from the semiannular sulcus (S sa) to the rhinal sulcus (S rh). The regions of the hippocampal formation are covered by an allocortex. The allocortex has fewer layers than the six-layered isocortex. A closer look at the entorhinal cortex seems to contradict this rule, as Braak identified up to 12 entorhinal layers [9, 10]. However, ontogenetic studies show that these sublayers arise from three-layered precursors [11]. Large chromophilic stellate or pre-alpha cells are specific cells of the entorhinal region. They are arranged in clusters and constitute the second layer of the entorhinal cortex. As a rule, the pre-alpha clusters are replaced by a chromophilic layer II of small cells in the medial wall of the rhinal sulcus. This feature marks the transentorhinal region, a region of transition between allo-and isocortex.
Figure 2b differs from Fig. 2a by a general decrease in staining intensity or pallor. This is evident when comparing Ch4p, the chromophilic neurons in the amygdaloid nuclei, and the entorhinal pre-alpha cells. In addition, Ce, Me, BM, and BL show sectors with significant neuronal loss along with thinning of layers in the hippocampal formation.
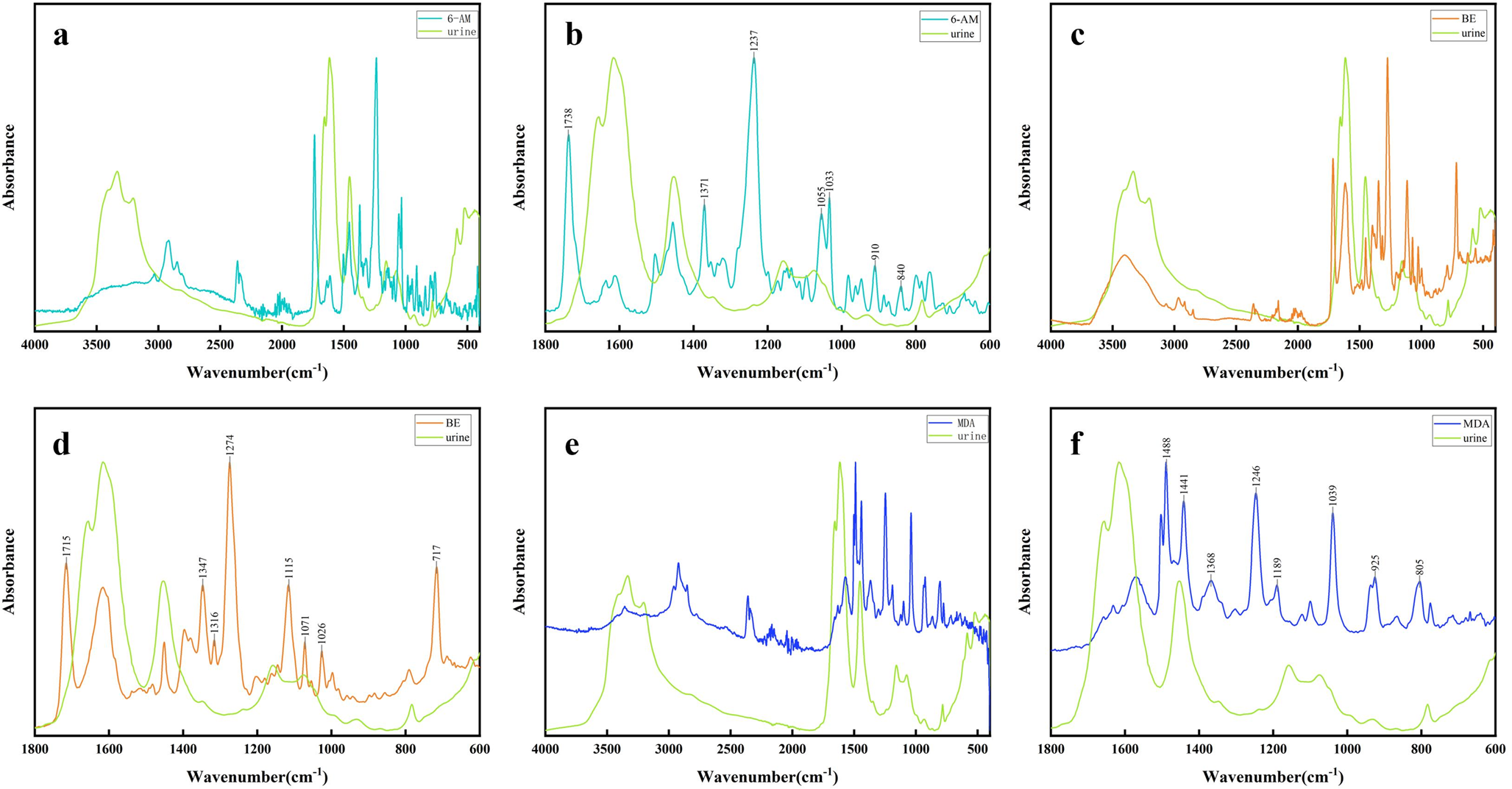
The two types of neurons are distinguished by their size, shape, and radiating bi-multipolar dendritic processes. Nissl bodies accumulate at the base of the dendritic arbor of pre-alpha cells (Fig. 3a, arrows) and the soma of large Ch4p cells (Fig. 3c and d arrows). A further distinguishing feature of pre-alpha cells is the greater chromaticity of their perikaryon compared to that of large Ch4p cells. The presence of nucleoli was observed in both cell types, while the nuclear membrane was only barely discernible. We have inserted the inset in Fig. 3c to visualize the amount and distribution of Nissl bodies in 87-year-old female control case conventionally fixed with formalin. The arrow in the inset of Fig. 3c points to a lobated nucleus with two nucleoli. The central parts of large basal Meynert neurons are filled with a light yellow to brown material, lipofuscin, which is known to increase with age and to dislocate Nissl bodies to peripheral perikaryal (cytoplasmatic) compartments of the nucleus basalis Meynert neurons (Fig. 3d arrows). After formalin fixation Nissl bodies are more numerous, their variation in size and distribution can be better assessed after formalin fixation compared to brain tissue subjected to heat exposure (Fig. 3c and d).
Fig. 3
a) and b) entorhinal pre-alpha cells, c) and d) large projection neurons of Meynert’s basal nucleus Ch4 p. a and c from Case 1, b and d from Case 2. Insetc 87-year-old woman, control case (vaginal cancer as cause of death)
The nuclear staining of glial cells contrasts with that of neurons. The nuclear membrane and heterochromatin are clearly delineated (Fig. 3c and d blockface arrows). Astrocytes are easily diagnosed; the differential diagnosis of oligodendrocytes and microglial cells is uncertain.
The neuron density of pre-alpha cells in the entorhinal region of the 77-year-old male subject is lower compared with the identical region in the 44-year-old male subject. Conversely, the glial cell density exhibited a conspicuous enhancement in the older individual (Fig. 3a and b).
Cytoarchitectonic studies require low-power magnification and strictly perpendicular sections through defined regions of the gyri under study (crown, wall, fundus). Cytoarchitectonic features of isocortical fields are intermediate between agranular (motor) and granular (koniocortical) sensory fields. They consist of 6 layers designated by Roman numbers, beginning with the superficial layer I and ending with the deep layer VI. Layers III, V, and VI are subdivided into sublayers termed a, b, and c [12].
The size, shape, orientation, and density of neurons in the cortical layers, as well as their relative thickness, provide criteria for delineating cortical fields. Layers I through VI are stacked horizontally, however, with higher magnification fibers are seen to leave the central medullary ray of the gyri and arch vertically through the cortical layers. These radial fibers cause a region-specific cortical striation and vertical columnar organization of neurons. The number and thickness of radial fibers provide additional criteria for delineating architectural fields. Rectangles in Fig. 1a and b illustrate the topography in the whole slice.
Low-power magnification reveals significant differences between the two cases under investigation, including pallor of the supragranular layers III and loss of layer IV granule cells. The presence of intensely stained layer Va pyramidal cells and neuron loss in layers Vb and VI create a rudimentary horizontal layering, which is already recognizable in the overview panel (Figs. 1b and 4b). The remaining radial fibers appear less numerous and thicker.
Another striking finding is the markedly increased chromophilia of the central medullary ray in the 77-year-old male subject. The alterations in stainability are not confined to the central medullary ray; they are also present in the central gray matter (Fig. 1b).
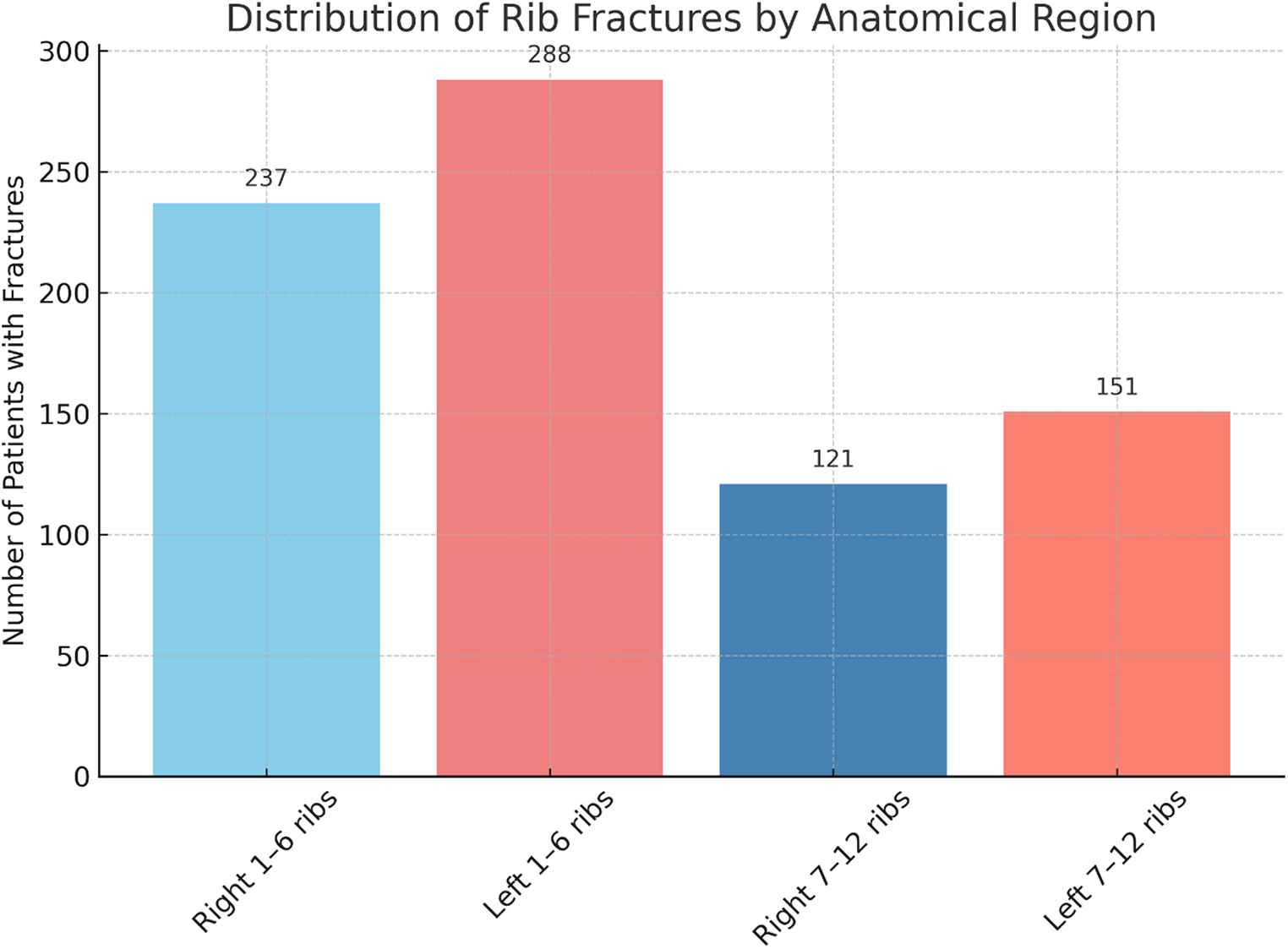
Pyramidal cells constitute approximately 80% of all telencephalic cortical neurons [13]. The rectangles in Fig. 4a and b demarcate the position of neurons and glial cells. The staining intensity of neurons, glial cells, and neuropil (diffusely stained background consisting mainly of abundant fine invisible neurites and glial processes) is higher in the 44-year-old male subject. Nissl bodies (or better substance) are rare in IIIa pyramidal cells (arrows in Fig. 4c) and in IIIc pyramidal cells (arrow in Fig. 4e).
The perikaryon of IIIa and IIIc-pyramidal cells is pale in area 22 of the 77-year-old male subject. Only one Nissl body could be identified in a IIIc pyramidal cell (arrow in Fig. 4f). Prominent nuclei and nucleoli contrast with the pale surrounding perikaryon.
The inset in Fig. 4f shows a Nissl body and pyramidal cell dendrites (arrow), lipofuscin granules in the basal parts of a large IIIc pyramidal cell (Fig. 4f, inset, star), fine lipofuscin granules surrounding a reactive astrocyte (Fig. 4f, inset, block arrow) and yellow material engulfed by a microglial cell (Fig. 4f, inset, arrowhead).
The central medullary ray of human telencephalic gyri contains few, if any, neurons in its central parts (Fig. 4g block arrows). The nuclei of oligodendrocytes and far fewer nuclei of astrocytes (Fig. 4, g and h, black arrows) predominate in this part of the gyri. The density of oligodendrocytes is low in the 44-year-old male and very high in the 77-year-old male. The nuclei of oligodendrocytes can be arranged in long rows or in a semicircular fashion surrounding optically bright spaces with a single central astrocyte (Fig. 4h, black arrows).
Fig. 4
a) and c) superior temporal gyri at low power magnification; rectangles and letters indicate the location of figures c-h taken with an oil immersion microscopic lens; a, c, e, g Case 1, 44-year-old man; b, d, f, h Case 2, 77- year-old man. Inset f 87-year-old woman, control case (vaginal cancer as cause of death)
The coronal plane of the slices of our younger case passes through anterolateral parts of the anterior commissure (C a). The putamen (Put) is still connected to the caudate nucleus (Caud) by the ventral striatum. The anterior perforate substance ventral to Mesulams Ch4i forms the telencephalic floor in this region. It is perforated by the small anterolateral arteries arising from the M1 segment of the medial cerebral artery. These branches supply major parts of the external or internal pallidal globe (Gpe, Gpi), the putamen, parts of the internal capsule (C i) and the head of the caudate nucleus. The origin and course of the perforant arteries are subject to individual variation with different clinical importances [14].
The course of these branches is easy to follow in our thick sections because of their relatively wide lumina. The rectangle at the medial border of the pallidum with the lateral parts of the internal capsule encloses two segments of these arteries. We made optical sections at 40x microscopic magnification and combined the individual optical sections into a coherent 3D stack (inset in Fig. 5). The right-angled emergence of the arterioles is noteworthy because it exerts a lifelong hemodynamic stress on the arterial walls at these sites.
Fig. 5
Close-up view of the left basal ganglia and anterior perforate substance of Case 1, 44-year-old man; the inset shows stacked optical slices of two small crossing arteries in the internal pallidal globe
Comments (0)