
Remember me

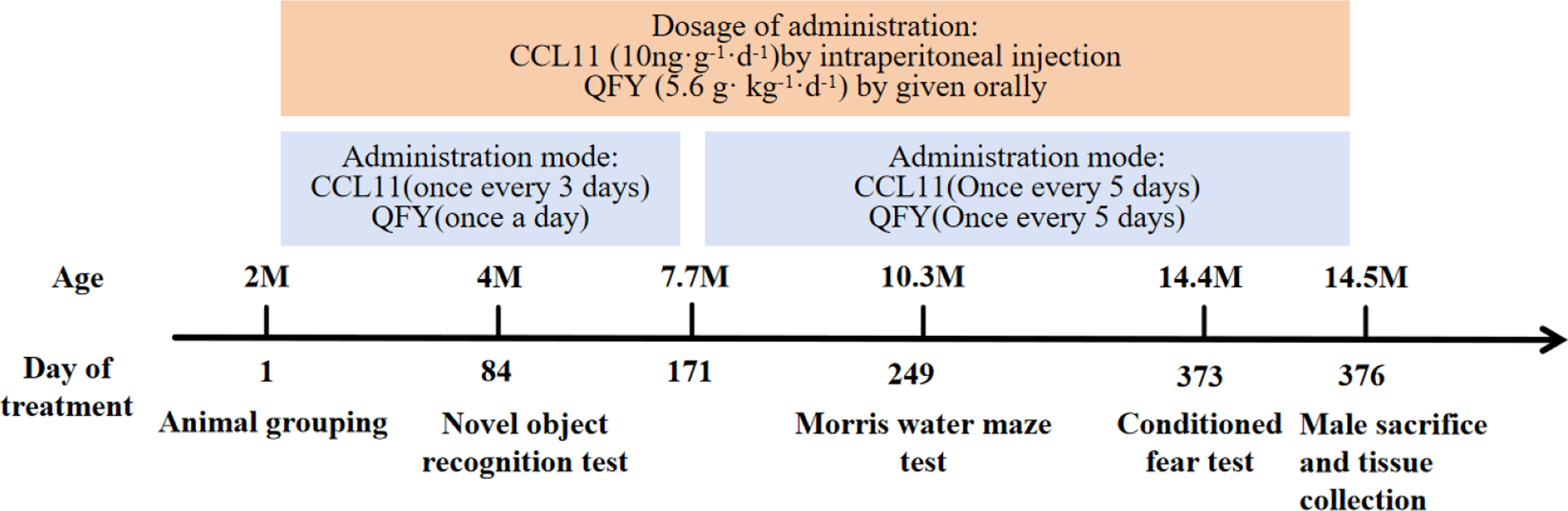

The novel object recognition test revealed that the preference index (PI) of mice following CCL11 intervention was significantly lower than that of normal C57BL/6 mice after a 24-hour learning period (Fig. 2 A-B, P < 0.05). Furthermore, the PI of mice treated with Qifuyin was significantly higher compared to that of mice subjected to CCL11 intervention (Fig. 2 A-B, P < 0.01). These results suggest that CCL11 injection impaired the object recognition memory function in 5-month-old male C57BL/6 mice, while Qifuyin significantly enhanced this cognitive index.
In the MWM test, CCL11 significantly increased the area under the curve (AUC) of escape latency during the learning period (Fig. 2 C-D, P < 0.05), reduced the number of platform crossings (Fig. 2E, P < 0.05), and increased the escape latency during the testing period (Fig. 2 F, P < 0.05). Following the administration of Qifuyin, the AUC of escape latency in the learning period was significantly reduced (Fig. 2 C-D, P < 0.05), the number of platform crossings was increased (Fig. 2E, P < 0.05), and escape latency during the testing period was significantly decreased (Fig. 2 F, P < 0.05).
In the fear conditioning test, the freezing time of C57BL/6 mice treated with CCL11 was significantly reduced compared to that of normal C57BL/6 mice (Fig. 2G, P < 0.01). Conversely, following the intervention with Qifuyin, the freezing time of mice during the test period was significantly prolonged (Fig. 2G, P < 0.01).
In summary, CCL11 appears to impair the object recognition ability, spatial learning and memory capacity, as well as situational fear memory of normal C57BL/6 mice. In contrast, Qifuyin demonstrates a protective effect on the cognitive abilities of mice against the damage induced by CCL11.
Fig. 2
Protective effects of Qifuyin on cognitive dysfunction in CCL11-induced C57BL/6 mice. (A-B): Novel objective recognition test. (A) Object preference index in learning period; (B) Test period, new object preference index after 24 h. (C-F): Morris water maze test. (C) Escape latency during the learning period; (D) Area under the curve of escape latency during the learning period; (E) Number of crossing the plat during the test period.; (F) Escape latency in the test period; G: Fear conditioning test. Freezing time of mice during the test period.Mean ± S.D., n = 8–13, *P#x2009;< 0.05, **P < 0.01 vs. Control; #P < 0.05, ##P < 0.01 vs. Model, Student’s t-test, analysis by GraphPad Prism 8.0.1
CCL11 induces synaptic plasticity impairment in C57BL/6 mice, while Qifuyin can restore synaptic plasticity in these miceNissl staining was employed to assess neuronal activity (Fig. 3 A). In comparison to normal C57BL/6 mice, the cell boundaries of hippocampal neurons in the model group appeared blurred, and the cytoplasm was less pronounced. Following treatment with Qifuyin, hippocampal neurons exhibited structural integrity, with neatly arranged multi-layer vertebral cells, dark blue-stained cytoplasm, and a high abundance of Ni-type corpuscles, in contrast to mice treated with CCL11. Statistical analysis indicated a significant reduction in neuronal activity in the model group compared to normal C57BL/6 mice (Fig. 3B, P < 0.01). Moreover, there was a significant increase in neuronal activity in the hippocampus of mice treated with Qifuyin compared to those subjected to CCL11 intervention (Fig. 3B, P < 0.05).
To evaluate the effects of Qifuyin on the synaptic plasticity of C57BL/6 mice induced by CCL11, we assessed the long-term potentiation (LTP) of the CA3-CA1 pathway in the hippocampus using MEA technology. Our findings indicate that CCL11 significantly inhibited the development of LTP in the CA3-CA1 pathway compared to control mice (Fig. 3 C-D, P < 0.01). Following treatment with Qifuyin, the slope of the field excitatory postsynaptic potential (f EPSP) was significantly greater than that observed in mice treated with CCL11 alone (Fig. 3 C-D, P < 0.01).
Compared to normal C57BL/6 mice, those treated with CCL11 exhibited a statistically significant decrease in dendritic spine density in neurons (Fig. 3 F-G, P < 0.01). Additionally, the number of junctions along the Shaw grid also significantly decreased (Fig. 3G-H, P < 0.01). In contrast, mice treated with Qifuyin demonstrated a substantial increase in the number of intersections along the Shaw grid (Fig. 3 H-I, P < 0.01) and a significant increase in dendritic spine density (Fig. 3 F-G, P < 0.01).
The synaptic structure of normal C57BL/6 mice was well-preserved, exhibiting distinct anterior and posterior membranes along with a significant quantity of presynaptic membrane vesicles, as determined by transmission electron microscopy. In contrast, mice treated with CCL11 showed a marked reduction in the number of intact synaptic structures in the hippocampus, characterized by a decrease in presynaptic membrane vesicles, lighter staining, and indistinct boundaries of the postsynaptic membrane. Notably, when comparing the Qifuyin group to the model group, the former exhibited an increased number of vesicles in the anterior membrane, intensified staining, clearly defined boundaries between the anterior and posterior membranes, and a higher density of substances in the posterior membrane (Fig. 3 J).
The expression levels of SYN, GAP-43, and PSD-95 in the hippocampus of C57BL/6 mice were assessed using ELISA. Compared to normal C57BL/6 mice, the levels of SYN (Fig. 3 K, P < 0.01), GAP-43 (Fig. 3 L, P < 0.01), and PSD-95 (Fig. 3M, P < 0.05) in the hippocampus of mice treated with CCL11 were significantly reduced. Following treatment with Qifuyin, the levels of these three proteins in the hippocampus of the mice exhibited a significant increase (Fig. 3 K-M, P < 0.01).
These findings suggest that CCL11 may impair neuronal activity in the hippocampus, disrupt synaptic structure, and inhibit LTP. Conversely, Qifuyin appears to protect hippocampal synaptic plasticity from damage induced by CCL11.
Fig. 3

Protective effect of QFY on impairment of synaptic plasticity induced by CCL11 in C57BL/6 mice. A. Nissl staining; B. Analysis of optical density values of Nissl staining(n = 3). C-D: LTP. C: EPSP slope of experimental groups; D: Amplitude of LTP in each group (n = 3). E-I: Golgi staining. E. Representative images showing Golgi staining of hippocampal brain tissue samples from mice in each group. (scale = 200 μm); F. Representative dendritic spine morphological map of neurons in the hippocampal CA1 regions from mice in each group. (scale = 5 μm); G. Quantitative analysis of the total dendritic spine density in the hippocampal CA1 regions of mice in each group. The data are presented as the mean ± SD (n = 9); H. Line chart of intersection points along Shaw grid; I. Area under Shaw grid line chart curve. J: Synaptic microstructure in mouse hippocampus. K-M: Expression of synapse-related proteins. K. SYN (n = 7–8); L. GAP-43 (n = 7–8); M. PSD-95 (n = 8).Mean ± S.D., Student`s t-test; *P < 0.05, **P < 0.01 compared to the Control group; #P < 0.05, ##P < 0.01 compared to the Model group, Student’s t-test, analysis by GraphPad Prism 8.0.1
CCL11 induces senescence in microglia, while Qifuyin can inhibit microglial senescenceFollowing labeling, the senescent cells were observed as irregular blue granules, as depicted in the image. In comparison to normal C57BL/6 mice, the number of SA-β-gal positive cells in the hippocampus of mice treated with CCL11 increased, while the number of SA-β-gal positive cells decreased after treatment with Qifuyin (Fig. 4 A).
Immunofluorescence results indicated that the quantity of p21 positive microglia was significantly higher than that in normal C57BL/6 mice (Fig. 4 C, P < 0.05). Furthermore, treatment with Qifuyin significantly reduced the expression of p21 in microglia (Fig. 4 C, P < 0.01).
Compared to normal C57BL/6 mice, plasma levels of INF-γ (Fig. 4D, P < 0.05), MCP-1 (Fig. 4E, P < 0.05), IL-1α (Fig. 4 F, P < 0.05), and CCL11 (Fig. 4G, P < 0.01) were significantly elevated. In contrast, plasma levels of MCP-1 (Fig. 4E, P < 0.05), INF-γ (Fig. 4D, P < 0.01), and CCL11 (Fig. 4G, P < 0.05) in mice treated with Qifuyin were significantly lower than those in mice subjected to CCL11 intervention.
In the CCL11 intervention group, the levels of CCL11 (Fig. 4 H, P < 0.001) and CCR3 (Fig. 4I, P < 0.05) in the brain were significantly increased, while the CCL11 levels (Fig. 4 H, P < 0.05) significantly decreased following Qifuyin treatment.
These results indicate that CCL11 can enhance the senescence-associated secretory phenotype (SASP) in plasma and increase the number of SA-β-gal positive cells in the brains of normal C57BL/6 mice. Additionally, the co-localization of the aging marker p21 with microglia suggests that CCL11 can induce microglial senescence. However, Qifuyin appears to protect microglia from senescence induced by CCL11.
Fig. 4

Qifuyin ameliorates microglia senescence induced by CCL11 in the brain of C57BL/6 mice. A: β-galactosidase staining in the hippocampus of mice. B. Co-location of p21 and Iba1; C. Quantitative of Iba1+p21+ cells. D-G: SASP content in plasma. D. INF-γ; E. MCP-1; F. IL-1α; G. CCL11. H. CCL11 content in the brain. I. CCR3 content in the hippocampus. Mean ± S.D., n = 3–8, *P < 0.05, **P < 0.01, ***P < 0.001 vs. Control; #P < 0.05, ##P < 0.01 vs. Model. Student’s t-test, analysis by GraphPad Prism 8.0.1
Effects Qifuyin on the senescence of CCR3-OE HMC3 cellsTo investigate whether CCL11 promotes microglial aging through its receptor CCR3, a stable cell line, CCR3-OE HMC3, was established for this experiment. Additionally, since Qifuyin is a traditional Chinese medicine compound consisting of components that are either not absorbed into the bloodstream, absorbed into the bloodstream, or able to cross the blood-brain barrier, this study utilized five monomer components that penetrate the brain as substitutes for Qifuyin, based on previous research conducted by our team.
Compared to the control group, the cell cycle in the sham group exhibited no significant changes (Fig. 5 A-O). In contrast, overexpression of CCR3 resulted in a notable increase in the proportion of HMC3 cells in the G0/G1 phase compared to the sham group (Fig. 5P, P < 0.001). Following the intervention with CCL11 on HMC3 cells overexpressing CCR3, a significant increase in the proportion of cells in the G0/G1 phase was observed (Fig. 5P, P < 0.05). Furthermore, compared to the CCR3-OE + CCL11 group, the proportion of cells in the G0/G1 phase was significantly reduced after intervention with SB297006 (Fig. 5P, P < 0.01), 3’,6-mustard acyl sucrose (Fig. 5P, P < 0.05), ginsenoside Rg1 (Fig. 5P, P < 0.05), and 18β-glycyrrhetinic acid (Fig. 5P, P < 0.05). Simultaneously, the inhibitors SB297006 (Fig. 5R, P < 0.05), 3’,6-mustard acyl sucrose (Fig. 5R, P < 0.05), and ginsenoside Rg1 (Fig. 5R, P < 0.01) significantly increased the proportion of cells in the S phase.
The expression of p16 (Fig. 5. T, P < 0.001) and p21 (Fig. 5. V, P < 0.001) in CCR3-OE HMC3 cells increased significantly in comparison to the sham group. The expression of p21 increased substantially (Fig. 5. V, P < 0.05) following the CCL11 intervention, while p16 did not exhibit any significant changes. The SB297006 and the five brain-entering components of Qifuyin can substantially reduce the expression of p16 and p21 in cells in comparison to the CCR3-OE + CCL11 group.
The expression levels of p16 (Fig. 5T, P < 0.001) and p21 (Fig. 5 V, P < 0.001) in CCR3-OE HMC3 cells were significantly elevated when compared to the sham group. Notably, the expression of p21 showed a substantial increase (Fig. 5 V, P < 0.05) following CCL11 treatment, whereas p16 did not demonstrate any significant alterations. Furthermore, both SB297006 and the five brain-penetrating components of Qifuyin were found to significantly reduce the expression levels of p16 and p21 in cells compared to the CCR3-OE + CCL11 group.
Fig. 5

Qifuyin reduce senescence induced by CCL11 in CCR3-OE HMC3 cell. A-R: Typical representative figures of cell cycle detection in each group by flow cytometry. A. Con: HMC3; B. Sham: HMC3-Sham; C. OE: HMC3 CCR3-OE; D. OE + CCL11:CCL11; E. SB: SB297006; F. C-DISS: 3’,6-Disinapoylsucrose; G. C-G-Rg1: Ginsenoside Rg1; H. C-TEN: Tenuifolin; I. C-18β-GA: 18β-Glycyrrhetinic acid; J. C-LQ: Liquiritin; K. DISS: 3’,6-Disinapoylsucrose; L. G-Rg1: Ginsenoside Rg1; M. TEN: Tenuifolin; N. 18β-GA: 18β-Glycyrrhetinic acid; O. LQ: Liquiritin; P. Effects of Qifuyin on G0/G1 phase of HMC3 CCR3-OE cells; Q. Effects of Qifuyin on G2/M phase of HMC3 CCR3-OE cells; R. Effects of Qifuyin on S. phase of HMC3 CCR3-OE cells. U-V. Expression of p16 in HMC3 CCR3-OE cells; P-Q: Expression of p21 in HMC3 CCR3-OE cells. Mean ± S.D., n = 3, ***P < 0.001 vs. Sham group, #P < 0.05, ##P < 0.01, ###P < 0.001 vs. CCR3-OE group, Student’s t-test; &P < 0.05, &&P < 0.01, &&&P < 0.001 vs. CCR3-OE + CCL11 group, One-way ANOVA analysis by GraphPad Prism 8.0.1
After 24 h of intervention with brain-entering components of Qifuyin on CCR3-OE HMC3 cells, the levels of IL-1β, INF-γ, IL-10, and TGF-β1 in the cell supernatant were measured using ELISA and Luminex technology. The results indicated that, compared to the sham group, the levels of the four cytokines in the cell supernatant of the CCR3-OE group were significantly elevated (Fig. 6 A-D, P < 0.05), with no significant changes observed following CCL11 intervention. In comparison to the CCR3-OE + CCL11 group, the levels of the four cytokines were significantly reduced in the SB, DISS, G-Rg1, and TEN groups (Fig. 6 A-D, P < 0.01). Additionally, 18β-GA was found to significantly decrease the levels of IL-1β (Fig. 6A, P < 0.05), INF-γ (Fig. 6B, P < 0.001), and TGF-β1 (Fig. 6D, P < 0.001). Furthermore, LQ significantly reduced the levels of IL-1β (Fig. 6 A, P < 0.01) and INF-γ (Fig. 6B, P < 0.001).
After CCL11 stimulation of HMC3 cells, which overexpress the CCR3 receptor, there was a significant increase in the proportion of cells in the G0/G1 phase. Additionally, the expression levels of aging markers p16 and p21 increased significantly, along with a notable rise in the content of the SASP. Furthermore, the senescent state of the cells was markedly improved following pretreatment with components of Qifuyin entering the brain.
Fig. 6
Qifuyin decrease SASP level in supernatant of HMC3 CCR3-OE cells. (A) IL-1β; (B) INF-γ; (C) LI-10; (D) TGF-β1. Control: HMC3; Sham: sham HMC3; OE: CCR3-OE HMC3; OE + CCL11:CCL11; SB: SB297006; DISS: 3’,6-Disinapoylsucrose; G-Rg1: Ginsenoside Rg1; TEN: Tenuifolin; 18β-GA: 18β-Glycyrrhetinic acid; LQ: Liquiritin. Mean ± S.D., n = 4–8, *P < 0.05, **P < 0.01 vs. Sham group, Student’s t-test; &P < 0.01, &&P < 0.05, &&&P < 0.001 vs. OE + CCL11 group, One-way ANOVA analysis by GraphPad Prism 8.0.1
Protective effect of the brain-entering components in Qifuyin on the neurite growth of HT-22 cells affected by the conditional medium of CCR3-OE HMC3To further investigate whether CCL11-induced senescence of microglia can impair the synaptic plasticity of neurons, we employed an indirect co-culture system in vitro. This approach allowed us to observe the effects of CCR3-O3 HTMC3 cell supernatant, treated with CCL11 and Qifuyin, on neurite formation in HT-22 cells.
Upon activation by CCL11, HMC3 cells overexpressing CCR3 exhibit pronounced senescence, accompanied by a significant increase in the content of SASP in the cell supernatant. Following treatment with the supernatant from CCR3-overexpressing HMC3 cells, the process of HT-22 cells was markedly reduced compared to the sham operation group. Notably, in comparison to the OE-CCL11 group, the intervention of various brain groups with Qifuyin resulted in an increase in cell processes (Fig. 7 A). Furthermore, an ELISA was conducted to assess the expression of synapse-related proteins in HT-22 cells treated with the conditioned media from CCR3-overexpressing HMC3 cells that had been subjected to CCL11 and Qifuyin intervention. PSD-95 (Fig. 7B, P < 0.001), GAP-43 (Fig. 7 C, P < 0.001), and SYN1 (Fig. 7D, P < 0.001) expressions were considerably lower in the CCR3-OE group than in the Sham group. PSD-95 expressions were considerably decreased (Fig. 7B, P < 0.05) following CCL11 intervention compared to the CCR3-OE group. When compared with CCR3-OE + CCL11 group, the expression of PSD-95 in SB (Fig. 7B, P < 0.05), G-Rg1 (Fig. 7B, P < 0.001), TEN (Fig. 7B, P < 0.05), 18β-GA (Fig. 7B, P < 0.05) and LQ (Fig. 7B, P < 0.01) groups was significantly higher. The expression of GAP-43 in SB (Fig. 7 C, P < 0.001), G-Rg1 (Fig. 7 C, P < 0.01) and 18β-GA (Fig. 7 C, P < 0.001) groups increased significantly. Only G-Rg1 (Fig. 7D, P < 0.05) and TEN (Fig. 7D, P < 0.05) can significantly increase the expression of SYN1 in HT-22 cells.
After stimulation by CCL11, HMC3 cells overexpressing CCR3 exhibit a significant growth restriction of HT-22 cells, accompanied by a reduction in the expression of synapse-related proteins, including SYN, GAP-43, and PSD-95. However, the extent of damage inflicted by CCR3-overexpressing HMC3 cells on HT-22 cell processes was notably diminished following pre-protection with the brain components of Qifuyin.
Fig. 7
Protective effect of Qifuyin on neural outgrowth of HT-22 cells injured by HMC3 CCR3-OE conditioned medium. A: The Cell morphology of HT-22. B-D: The expression of synapse related proteins in HT-22 cells. B. PSD-95; C. GAP-43; D. SYN1.Mean ± S.D., n = 4–8, ***P < 0.001 vs. Sham group, #P < 0.001 vs. CCR3-OE group, Student’s t-test; &P < 0.01, &&P < 0.05, &&&P < 0.001 vs. OE + CCL11 group, One-way ANOVA analysis by GraphPad Prism 8.0.1
Effects Qifuyin on the senescence of CCR3-KO HMC3 cellsIn order to verify whether CCL11 promotes microglia senescence by activating CCR3 receptor, we constructed HMC3 cells with CCR3 receptor knocked out, and observed the cell cycle by flow cytometry and the expression of senescence markers by IF.
In comparison to the Con group, the sham group exhibited no significant changes in the cell cycle. Notably, the CCR3-KO group showed a significant increase in the proportion of cells in the G0/G1 phase when compared to the sham group (Fig. 8 K, P < 0.05). Furthermore, there were no significant changes in the cell cycle of the CCR3-KO group after CCL11 intervention. In contrast to the CCR3-KO + CCL11 group, the proportion of cells in the G0/G1 phase was significantly decreased following treatment with DISS (Fig. 8 K, P < 0.05) or G-Rg1 (Fig. 8 K, P < 0.01). Additionally, the proportion of cells in the S phase of the G-Rg1 group was significantly increased (Fig. 8M, P < 0.05).
The expression levels of p16 (Fig. 8 N) and p21 (Fig. 8P) in CCR3-KO HMC3 cells were evaluated using IF following treatment with five brain-entering components, specifically CCL11 and Qifuyin. The results indicated that, compared to the Sham group, the expression of p16 (Fig. 8O, P < 0.05) and p21 (Fig. 8Q, P < 0.05) in CCR3-KO HMC3 cells was significantly decreased (P < 0.05). In contrast, the other compounds did not exhibit a significant effect on the expression levels of p16 and p21.
The results indicate that the reduction of CCR3 receptor expression on the cell surface effectively inhibits cellular senescence. Even upon re-administration of CCL11, it fails to induce cell senescence, demonstrating that CCL11 can only promote microglial activity following its binding to the CCR3 receptor.
Fig. 8

Effects of brain components of Qifuyin on the senescence of CCR3-KO HMC3 cells. A-J. Typical representative figures of cell cycle detection in each group by flow cytometry. (A) Con: HMC3; (B) Sham: HMC3-Sham; (C) KO: HMC3 CCR3-KO; (D) KO + CCL11:CCL11; E. SB: SB297006; F. DISS: 3’,6-Disinapoylsucrose; G. G-Rg1: Ginsenoside Rg1; H. TEN: Tenuifolin; I. 18β-GA: 18β-Glycyrrhetinic acid; J. LQ: Liquiritin; K. Effects of Qifuyin on G0/G1 phase of CCR3-KO HMC3 cells; L. Effects of Qifuyin on G2/M phase of CCR3-KO HMC3 cells; M. Effects of Qifuyin on S phase of CCR3-KO HMC3 cells. N-O. Expression of p16 in CCR3-KO HMC3 cells; P-Q: Expression of p21 in CCR3-KO HMC3 cells. Mean ± S.D., n = 3, *P < 0.05 vs. Sham group, #P < 0.05, ##P < 0.01 vs. KO + CCL11 group, Student’s t-test, analysis by GraphPad Prism 8.0.1
Comments (0)