
Remember me





In this study, we used iPSCs from four healthy individuals and four PD patients harboring the LRRK2-G2019S mutation (Table 1). Initially, embryoid bodies were formed from the iPSCs and cultured in a flask to produce macrophage precursors (Fig. 1A). These precursors were harvested and differentiated for 14 days to generate mature microglia, according to the previously established protocol [17, 24]. Healthy iPSC derived microglia expressed typical macrophage and microglial markers such as IBA1 and PU1 (Fig. 1B). After validating our robust microglial generation protocol, we compared microglia from healthy individuals and those with the LRRK-G2019S mutation.
Fig. 1
Generation and characterization of iPSC-derived microglia identity show no alteration in microglia harboring the LRRK2-G2019S mutation. A Schematic representation of the generation of iPSC-derived microglia. B Representative immunostaining image of healthy iPSC-derived microglia with Hoechst, PU1, and IBA1. C Flow cytometry analysis of LRRK2-WT and LRRK2-G2019S microglia viability upon differentiation using the Zombie NIR™ Fixable viability kit. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials D Luminescence-based quantification of cellular ATP levels in LRRK2-WT and LRRK2-G2019S microglia. Data are shown mean ± SD (n = 3–4), pooled from three independent trials. E Representative immunostaining images of LRRK2-WT and LRRK2-G2019S microglia with Hoechst, PU1, IBA1 (left) and Hoechst, CD11b, CD45 (right). F Flow cytometry analysis of mature microglia (CD11bhiCD45int) (left) and representative analysis of mean fluorescence intensity of CD11b and CD45 (right) in LRRK2-WT and LRRK2-G2019S microglia. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. G. Representative analysis of mean fluorescence intensity of microglia identity markers (P2RY12, IBA1, TMEM119, CX3CR1) in mature LRRK2-WT vs LRRK2-G2019S microglia using flow cytometry
Despite reports of LRRK2-related toxicity resulting in 10–70% cell death, other studies found no effect on cell viability [25]. In our system, we observed no differences in cell viability as assessed by flow cytometry (Fig. 1C), or total ATP levels using CellTiter-GLO® luminescence assay (Fig. 1D). Additionally, the presence of the LRRK2-G2019S mutation did not impact embryoid body formation or the yield of macrophage precursors (Figure S1A). Furthermore, morphological assessments via flow cytometry (FSC/SSC) revealed no differences in cell size or complexity between healthy and mutant microglia (Figure S1B).
Upon stimulation IL-34 and GM-CSF, macrophage precursors mature into microglial cells [24]. Mature microglia express markers, including general macrophage markers (CD11b, CD45, IBA1, PU1) as well as specific microglial markers (P2RY12, CX3CR1, and TMEM119) [26]. Both wild-type (WT) and LRRK2-G2019S microglia showed comparable expressions of these markers, as confirmed by immunofluorescence and flow cytometry (Fig. 1E-G, S1C). Together, these data indicate that the LRRK2-G2019S mutation does not alter the differentiation efficiency, identity, or basic morphology of iPSC-derived microglia.
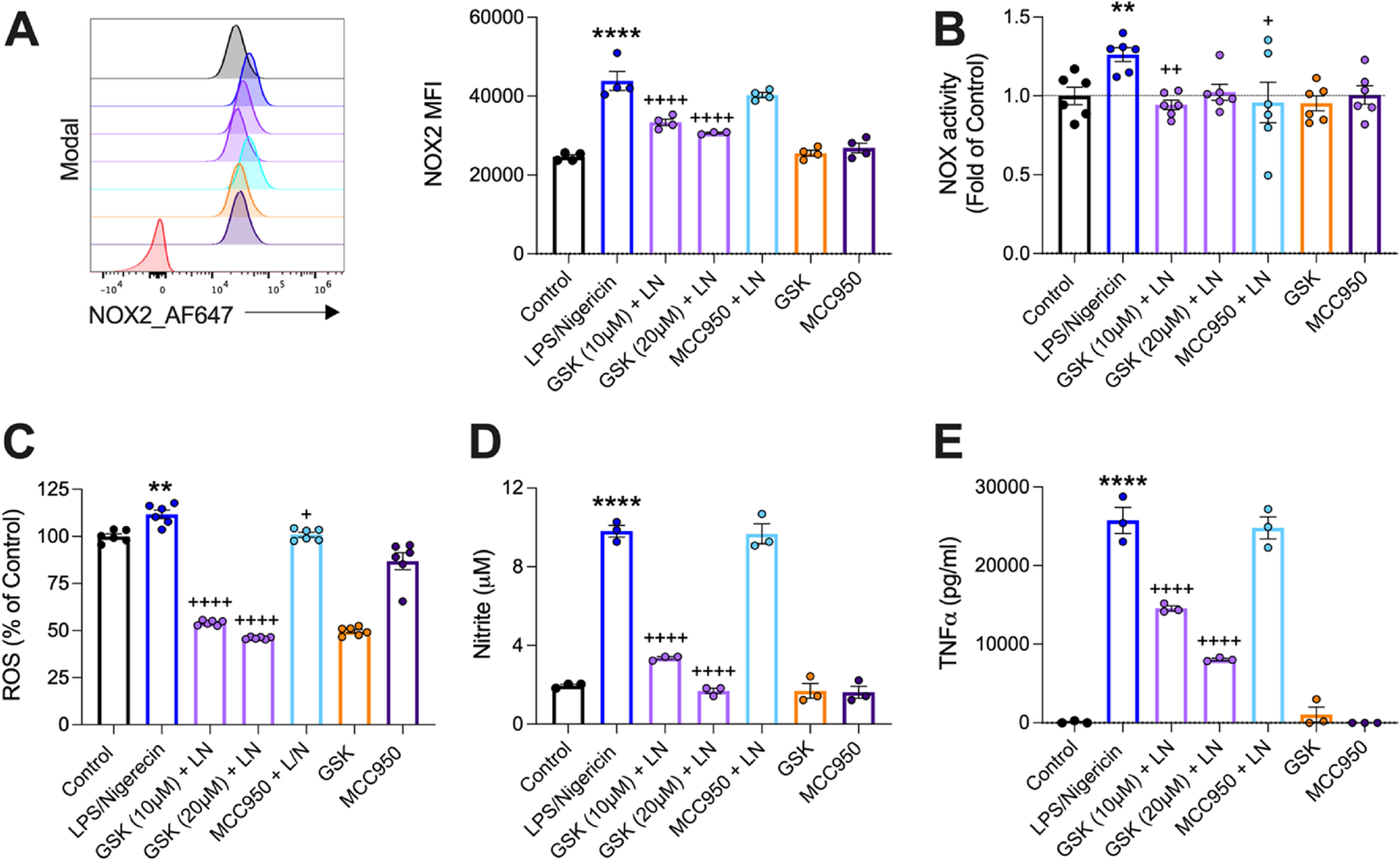
LRRK2-G2019S microglia exhibit heightened immune activation and inflammatory phenotypesDespite normal differentiation, LRRK2-G2019S microglia exhibit increased immune activation and proinflammatory properties. Previous studies have implicated LRRK2 polymorphisms in autoinflammatory diseases including inflammatory bowel disease (IBD), Crohn’s disease, and tuberculosis, indicating a significant connection to immune functions [27]. Notably, increased expression of LRRK2 has been observed in immune cells in response to proinflammatory signals [28]. We therefore sought to determine the impact of LRRK2-G2019S mutation on microglial inflammatory functions in iPSC-derived microglia.
Flow cytometry analysis showed that CD68, a lysosomal protein indicative of phagocytic activity, was significantly upregulated in LRRK2-G2019S microglia compared to healthy controls (Fig. 2A). Markers associated with antigen presentation and costimulation, including HLA-DR, CD80 and CD86, were elevated in mutant microglia relative to that in control microglia (Fig. 2A). However, other activation markers such as CD69, CCR6, and the exhaustion marker PD-1 were unchanged (Figure S2A).
Fig. 2
Immunological phenotyping of LRRK2-G2019S microglia showed heightened activation and inflammatory profiles. A Representative analysis of mean fluorescence intensity (top) and the quantification (bottom) of activation markers (CD68, HLA-DR, CD80, and CD86) of LRRK2-WT and LRRK2-G2019S microglia. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. B Representative analysis of mean fluorescence intensity (left) and the quantification (right) of pathogen recognition receptors TLR2 and TLR4 in LRRK2-WT and LRRK2-G2019S microglia. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. C Flow cytometry analysis of microglia phagocytosis in LRRK2-WT and LRRK2-G2019S microglia using fluorescently labelled Zymosan beads. Representative images are shown on the left, quantification on the right. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. D Flow cytometry analysis of microglia phagocytosis in LRRK2-WT and LRRK2-G2019S microglia using pH-sensitive fluorescent Zymosan beads. Representative images are shown on the left, quantification on the right. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. E Enzyme-linked immunosorbent assay of human inflammatory cytokines in LRRK2-WT and LRRK2-G2019S microglia using the Human Inflammatory Cytokine Multiplex ELISA Kit from Arigo Biolaboratories. Data are shown as mean ± SD (n = 4), representative of two independent trials. F Flow cytometry analysis of intracellular TNF-α in LRRK2-WT and LRRK2-G2019S microglia upon 24 h stimulation with LPS. Representative images are shown on the left, quantification on the right. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
One of the key functions of microglia is the uptake of antigens for removal or presentation. Microglia possess several pathogen recognition receptors, namely Toll-like receptors (TLRs). TLR signaling and phagocytosis are hallmarks of the microglia-mediated immune response to infections [29]. The activation of TLR2 has been associated with neuronal injury [30]. Our data revealed that LRRK2-G2019S microglia presented increased levels of both TLR2 and TLR4, suggesting a potential increase in phagocytic activity (Fig. 2B). To assess microglial phagocytosis, we used fluorescence labelled Zymosan, a TLR2 agonist derived from yeast. Following incubation and thorough washing to remove unbound particles, LRRK2-G2019S microglia showed significantly greater uptake of zymosan than healthy microglia did (Fig. 2C). Using a more advanced fluorescence Zymosan particle coupled with a pH-sensitive pHRodo, which fluoresces only when the particle is digested in the lysosome, we confirmed an increased phagocytic rate in LRRK2-G2019S microglia (Fig. 2D).
Another critical function of microglia is the production of cytokines. In response to environmental cues, microglia can produce a range of pro- and anti-inflammatory cytokines. Proinflammatory cytokines such as IL-1, IL-6, IFN, and TNF-α have been implicated in the pathogenesis of PD [31]. Our results indicated that various inflammatory cytokines were similarly expressed between healthy and LRRK2-G2019S microglia. However, upon LPS stimulation, LRRK2-G2019S microglia uniquely secreted higher levels of TNF-α into the culture media than the control microglia (Fig. 2E). This finding was corroborated by flow cytometry, which revealed significant upregulation of intracellular TNF-α in LRRK2-G2019S microglia (Fig. 2F, Figure S2B). Interestingly, other inflammatory cytokines were expressed at similar levels in both groups (Fig. 2E, Figure S2B). In summary, our data suggest that the LRRK2-G2019S mutation enhances the inflammatory activity and function of microglia, which is primarily mediated by increased TNF-α expression.
Transcriptomic profiling reveals altered inflammatory profiles and ROS metabolism in LRRK2-G2019S microgliaTo gain a deeper understanding of the molecular changes induced by the LRRK2-G2019S mutation in microglia, we performed a bulk RNA sequencing analysis. We included four healthy controls and four iPSC-derived microglia carrying the LRRK2-G2019S mutation. The transcriptomic data identified over 970 genes whose expression significantly differed between control and mutant microglia (Fig. 3A, B). Pathway analysis revealed that several inflammatory signaling pathways were differentially regulated in LRRK2-G2019S microglia compared with healthy controls. Among these, TNF and IL-1 signaling, both of which are critical mediators of immune responses and inflammation, were notably altered (Fig. 3C).
Fig. 3
Transcriptomic profiling of LRRK2-G2019S microglia revealed more pro-inflammatory microglia phenotypes. A Volcano plot showing all genes with genes having log2FC > 1 and p-value < 0.05 highlighted, indicating differentially expressed genes between LRRK2-WT and LRRK2-G2019S microglia. B Top 50 differentially expressed genes (DEGs) between LRRK2-WT and LRRK2-G2019S microglia with the lowest p value. C Enrichment analysis using Metacore, showing top pathway maps involved in immunity. D Fold changes of genes from the GO term of innate and adaptive immunity with log2FC > 0.5. E Metacore enrichment analysis of pathways associated with cellular metabolism. F Heatmap of mTOR-regulated genes distinguishing LRRK2-WT from LRRK2-G2019S microglia upon hierarchical clustering
Our transcriptomic analysis also revealed distinct alterations in genes related to microglial inflammatory processes such as phagocytosis and inflammation (Fig. 3C). Genes involved in pathogen recognition and phagocytosis including TLR2, TLR4, TLR8, TLR9, and TLR10 – exhibited altered expression in LRRK2-G2019S microglia. Similarly, the expression of ZAP70, a key molecule involved in immune synapse formation, was altered (Fig. 3D, Figure S3A). Additionally, genes involved in antigen presentation, such as HLA-DRB5, HLA-DOB, and CD209 (DC-SIGN) also displayed increased expression in mutant microglia (Fig. 3D, Figure S3A). These findings highlight the broad impact of the LRRK2-G2019S mutation on microglial profiles.
Given the observed upregulation of inflammatory markers at both the RNA and protein levels in LRRK2-G2019S microglia, we further investigated the underlying cellular mechanism contributing to this heightened immune response. Recent studies have emphasized the pivotal role of immune metabolism in regulating cell development, proliferation, and function. Immune cells dynamically rewire their metabolic preferences in response to environmental cues and functional demands. We hypothesized that the LRRK2-G2019S mutation might impair microglial metabolism, contributing to their abnormal immune function.
Enrichment analysis identified several altered pathways in LRRK2-G2019S microglia, many of which were related to metabolism (Supplemental information Table 1). Notably, pathways such as reactive oxygen species (ROS) metabolism and mammalian target of rapamycin complex 1 (mTORC1) signaling were enriched in mutant microglia (Fig. 3E). mTORC1, which plays a key role in regulating cellular metabolism and immune responses, has been implicated in microglial functions [32, 33]. Similarly, our data revealed that genes regulating mTORC1 activity, including TSC1, TSC2, and RPTOR, were differentially expressed in LRRK2-G2019S microglia (Fig. 3F).
To gain a more comprehensive view of the metabolic changes associated with the LRRK2-G2019S mutation, we constructed context-specific metabolic models by integrating the RNA sequencing data with the Recon3 generic human metabolic model, using the rFASTCORMICS pipeline [20]. This model predicted significant alterations in several metabolic pathways, including energy metabolism, sugar metabolism, amino acid metabolism, and lipid metabolism. Importantly, the mutant microglia exhibited unique metabolic reactions that were absent in the healthy controls (Figure S3B). Further analysis identified the most dysregulated metabolic subsystems in LRRK2-G2019S microglia. These included ROS metabolism (particularly glutathione metabolism), lipid metabolism and glycolysis, all of which play key roles in maintaining cellular homeostasis and responding to stress (Figure S3C). Dysregulation of these metabolic pathways is likely to contribute to the pathological phenotypes observed in LRRK2-G2019S microglia.
LRRK2-G2019S microglia upregulate glycolysis and downregulate glucose-derived serine synthesisIn our previous analysis, transcriptomic profiling and metabolic modelling showed changes in microglia harboring the LRRK2-G2019S mutation. mTOR plays a crucial role in integrating extracellular and intracellular signals to regulate cellular metabolism and growth [34]. Given the observed changes in metabolic homeostasis at the transcriptomic level, we next sought to determine the functional status of mTOR by measuring its phosphorylation using flow cytometry. Interestingly, the phosphorylation of mTOR was comparable in LRRK2-G2019S microglia (Figure S4A). However, we detected a significant increase in the expression of phosphorylated S6 (pS6), a key downstream target of mTORC1 in LRRK2-G2019S microglia compared with that in healthy controls (Fig. 4A).
Fig. 4
Metabolic analysis indicated metabolic reprogramming in LRRK2-G2019S microglia. A Flow cytometry analysis of protein S6 phosphorylation in LRRK2-WT and LRRK2-G2019S microglia. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. B Measurement of oxygen consumption rate (OCR) in LRRK2-WT and LRRK2-G2019S microglia using the Seahorse Cell Mitochondrial Stress Test. Maximum respiration and spare respiratory capacity were quantified (right). Data are shown as mean ± SD (n = 3–4), representative from three independent trials. C Flow cytometry analysis of respiring mitochondria, calculated by the ratio between Mitotracker Deep Red™ (marking active mitochondria) and Mitotracker Green ™ (marking total mitochondria). Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. D Flux map of carbon metabolism (left) and mass isotopomer distribution of TCA cycle metabolites in LRRK2-WT and LRRK2-G2019S microglia incubated with U-13C6-glucose during the final 24 h of differentiation. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. E Measurement of extracellular acidification rate (ECAR) in LRRK2-WT and LRRK2-G2019S microglia using Seahorse Glycolysis Stress Test. Basal glycolysis and glycolytic capacity were quantified (right). Data are shown as mean ± SD (n = 3–4), representative from three independent trials. F Flow cytometry analysis of glucose uptake rate in LRRK2-WT and LRRK2-G2019S microglia using fluorescent glucose 2-NBDG. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. G Mass isotopomer distribution of pyruvate (M + 3) and lactate (M + 3) in LRRK2-WT and LRRK2-G2019S microglia incubated with U-13C6-glucose during the final 24 h of differentiation. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. H Transcript count of serine synthesis genes PHGDH, PSAT1, and PSPH in LRRK2-WT and LRRK2-G2019S microglia. I Flow cytometry analysis of intracellular protein PHGDH in LRRK2-WT and LRRK2-G2019S microglia. Data are shown as mean ± SD (n = 3–4), pooled from four independent trials. J Mass isotopomer distribution of serine synthesis (M + 3) in LRRK2-WT and LRRK2-G2019S microglia incubated with U-.13C6-glucose during the final 24 h of differentiation. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
Since mTORC1 signaling is known to regulate major metabolic pathways, such as mitochondrial respiration and glycolysis, the upregulation of mTORC1 in LRRK2-G2019S microglia could impact these processes. To assess mitochondrial function, we performed a mitochondrial stress test using Seahorse flux analysis. Although the spare respiratory capacity remained similar, the maximum respiration rate as well as proton leak was significantly lower in LRRK2-G2019S microglia (Fig. 4B, SF4B).
Interestingly, the ratio of mitochondria with intact membrane potential (stained with MitoTracker DeepRed ™) to total mitochondrial mass (stained with MitoTracker Green™) was comparable between the mutant and control microglia, suggesting that mitochondrial quantity and membrane integrity were not compromised (Fig. 4C). Furthermore, the level of Ki-67 staining, a marker for cell proliferation, showed no differences between LRRK2-G2019S and healthy microglia (Figure S4C). Similarly, total ROS levels, as assessed using H2-DCFDA probe (Figure S4D), as well as mitochondria ROS (data not shown), were also comparable between the two groups. Normal ROS levels suggest that LRRK2-G2019S microglia do not experience excessive oxidative stress, despite their heightened metabolism and immune activity. These results indicate that mitochondrial function is largely preserved in LRRK2-G2019S microglia, and that the reduced reliance on mitochondrial respiration suggests a shift toward alternative energy pathways to sustain microglial activity.
To further explore the altered metabolic flux in LRRK2-G2019S microglia, we performed metabolic tracing using uniformly labelled 13C6 glucose. This approach allowed us to track the fate of glucose through various metabolic pathways. Pyruvate, the primary end product of glycolysis, can be used in the mitochondria to fuel the tricarboxylic acid (TCA) cycle. The mass isotopologue distribution (MID) showed no significant differences in the levels of TCA cycle intermediates – such as citrate, α-ketoglutarate, succinate, fumarate, and malate – between healthy and LRRK2-G2019S microglia (Fig. 4D). These findings align with the mitochondrial respiration results, further highlighting that the LRRK2-G2019S mutation does not significantly affect mitochondrial metabolism in microglia.
In resting immune cells, including microglia, energy production is typically reliant on mitochondrial metabolism; however, upon activation, these cells rewire their metabolism into glycolysis, a faster but less efficient metabolic pathway, to meet the increased energy demands [8]. Given the elevated immune activation in LRRK2-G2019S microglia, we suspected that glycolysis might be upregulated in these cells. To test this hypothesis, we performed a glycolytic stress test using Seahorse flux analysis. Indeed, basal glycolysis was elevated in LRRK2-G2019S microglia compared with healthy controls (Fig. 4E). However, their glycolytic capacity, defined as the maximum rate of glycolysis under stress conditions, was significantly reduced (Fig. 4E).
We further confirmed this metabolic shift by assessing glucose uptake using 2-NBDG, a fluorescent glucose analog. Compared with healthy microglia, LRRK2-G2019S microglia exhibited significantly greater glucose uptake (Fig. 4F). Instead of entering the TCA cycle, a significant portion of pyruvate generated from glycolysis was converted into lactate, as indicated by increased levels of M + 3 glucose-derived lactate (Fig. 4G). These data suggest that LRRK2-G2019S microglia undergo metabolic reprogramming and rely more on glycolysis to support their hyperactive immune function.
Glycolysis not only generates pyruvate but also produces intermediate metabolites that serve as precursors for other biosynthetic pathways. Specifically, 3-phosphoglycerate serves as a substrate for serine biosynthesis. Interestingly, two key genes involved in serine biosynthesis – PHGDH and PSAT1 – were significantly downregulated in LRRK2-G2019S microglia (Fig. 4H). PHGDH catalyzes the first and rate-limiting steps of serine synthesis. Flow cytometry confirmed a significant reduction in PHGDH expression in LRRK2-G2019S microglia compared with that in control microglia (Fig. 4I).
To further validate these findings, we traced the incorporation of glucose into serine using 13C6 glucose. The results revealed significantly lower levels of M + 3 glucose-derived serine in LRRK2-G2019S microglia than in control microglia, indicating an impaired serine biosynthetic pathway (Fig. 4J). The reduction in serine production may reflect a compensatory shift to drive enhanced glycolysis due to the LRRK2-G2019S mutation, which needs to be further elaborated. In summary, our data show that LRKR2-G2019S microglia exhibit metabolic reprogramming, characterized by elevated glycolysis and impaired serine synthesis, which might be targeted to restore normal microglial functions in PD.
LRRK2-G2019S microglia induce degeneration of dopaminergic neurons in midbrain organoidsNeuroinflammation has been implicated in the progressive degeneration of dopaminergic neurons in PD [35, 36]. Here, we demonstrated that LRRK2-G2019S microglia upregulate immune activity, which is characterized by increased phagocytosis and elevated production of the proinflammatory cytokine TNF-α. Given the established role of neuroinflammation in neuronal injury, we sought to explore how microglial immune activation contributes to dopaminergic neuron loss, with a specific focus on the pathological effects of the LRRK2-G2019S mutation.
To do so, we used midbrain organoids derived from healthy individual iPSCs [23]. To simulate the inflammatory conditions associated with LRRK2-G2019S microglia, we treated these healthy midbrain organoids with exogenous TNF-α. This cytokine exposure led to a significant reduction in the number of dopaminergic neurons, by up to 30% compared with that in untreated controls (Fig. 5A). Interestingly, in the co-culture of organoids with LRRK2-G2019S microglia, TNF-α neutralizing antibody rescued dopaminergic neuron degeneration to the WT level (SF4E). This result suggests that inflammatory signaling alone is sufficient to induce neuronal injury, mimicking aspects of LRRK2-G2019S microglial inflammation in a TNF-α-dependent manner and that blocking TNF-α protect dopaminergic neurons.
Fig. 5
LRRK2-G2019S microglia induced dopaminergic neuron degeneration in midbrain organoids. A The number of dopaminergic neurons, represented by the total TH + cells over total MAP2 + neurons, in midbrain organoids treated with or without 100 pg/ml TNF-α. Data are shown as mean ± SD (n = 3), pooled from three independent trials. B The number of dopaminergic neurons in midbrain organoids co-cultured with LRRK2-WT and LRRK2-G2019S microglia. Data are shown as mean ± SD (N = 3, n = 4), pooled from three independent trials. C Representative confocal image of a 70 μm midbrain organoids section with LRRK2-WT (top) or LRRK2-G2019S (bottom) microglia, immunostained with Hoechst, TH, MAP2, and IBA1. D High-content automated image analysis of immunofluorescence staining of dopaminergic neurons in assembloids (shown in C), expressed as the proportion of TH + cells normalized by total nuclei. Dotted color showed individual midbrain organoids. Data are shown as mean ± SD (N = 3, n = 4), pooled from three independent trials. p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
To further capture the complexity of the brain’s microenvironment and the contribution of microglia to dopaminergic neuron loss, we integrated iPSC-derived microglia into the midbrain organoids [17]. This coculture approach allowed us to assess the direct effects of both healthy and LRRK2-G2019S mutant microglia on neuronal health and survival. Flow cytometry analysis showed that healthy midbrain organoids cocultured with LRRK2-G2019S microglia had significantly lower levels of tyrosine hydroxylase (TH) positive cells, the rate-limiting enzyme in dopamine synthesis and a surrogate marker of dopaminergic neurons. In contrast, organoids cocultured with healthy control microglia retained higher levels of TH, suggesting that the mutant microglia had a neurotoxic effect on dopaminergic neurons (Fig. 5B).
This trend was further confirmed by imaging analysis. Immunostaining of the organoids revealed that the incorporation of both healthy and LRRK2-G2019S microglia was comparable. The total number of MAP2 neurons in both organoids were comparable. In contrast, the number of TH positive dopaminergic neurons was significantly reduced in LRRK2-G2019S microglia containing midbrain organoids (Fig. 5C, D). These data demonstrate that neuroinflammation driven by LRRK2-G2019S microglia is associated with reduced dopaminergic neuron markers in co-cultured midbrain organoids. Elevated levels of TNF-α produced by the mutant microglia appear to play a central role in this neurotoxicity.
Oxamic acid reduces immune activity and attenuates dopaminergic neuron lossThe finding that midbrain organoids cocultured with LRRK2-G2019S microglia experienced significant dopaminergic neuronal loss (Fig. 5B, C), indicates that microglia-mediated neuroinflammation plays a central role in PD pathogenesis. These results underscore the potential of targeting microglia as a strategy for disease modification. Given that impaired serine biosynthesis results in increased levels of mTOR and glycolysis, we speculated that targeting the metabolic dysregulation could mitigate neuroinflammation and prevent dopaminergic neuron loss.
To address this hypothesis, we aimed to target the serine-mTOR-glycolysis axis. We used oxamic acid, a pyruvate analog that inhibits lactate dehydrogenase by forming an inactive complex with the enzyme [37, 38]. Oxamic acid-mediated inhibition of lactate dehydrogenase has been shown to reduce glycolysis and mTOR-dependent metabolic reprogramming in cancer cells [39, 40]. Here, we aimed to determine whether targeting glycolysis in LRRK2-G2019S microglia would alleviate their inflammatory toxicity and rescue TH neuronal levels.
As predicted, compared with no treatment, oxamic acid significantly reduced glucose uptake in LRRK2-G2019S mutant microglia (Fig. 6A). This effect was selective, as glucose uptake in healthy microglia remained unaffected by the treatment, suggesting that oxamic acid specifically targets hyperactive glycolysis in LRRK2-G2019S microglia. Additionally, the elevated levels of pS6 in LRRK2-G2019S microglia were normalized to the control level upon treatment with oxamic acid (Fig. 6B).
Fig. 6
Targeting microglia metabolism with oxamic acid rescued LRRK2-G2019S microglia overt activation and preserved dopaminergic neurons in midbrain organoids. A Flow cytometry analysis glucose uptake rate using fluorescent glucose 2-NBDG in LRRK2-WT and LRRK2-G2019S microglia treated with or without oxamic acid. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. B Flow cytometry analysis of protein S6 phosphorylation in LRRK2-WT and LRRK2-G2019S microglia treated with or without oxamic acid. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. C Flow cytometry analysis of CD86 in LRRK2-WT and LRRK2-G2019S microglia treated with or without oxamic acid. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. D Flow cytometry analysis of microglia phagocytosis using pH-sensitive fluorescently labelled Zymosan beads in LRRK2-WT and LRRK2-G2019S microglia treated with or without oxamic acid. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. E Flow cytometry analysis of intracellular TNF-α in LRRK2-WT and LRRK2-G2019S microglia treated with or without oxamic acid. Data are shown as mean ± SD (n = 3–4), pooled from three independent trials. F Number of dopaminergic neurons in midbrain organoids co-cultured with LRRK2-WT and LRRK2-G2019S microglia treated with or without oxamic acid. Data are shown as mean ± SD (N = 3, n = 4), pooled from three independent trials
Next, we examined whether oxamic acid attenuate LRRK2-G2019S microglia immune hyperactivity. Flow cytometry analysis showed a significant reduction in the expression of the costimulatory molecules CD86 in LRRK2-G2019S microglia following oxamic acid treatment (Fig. 6C). Interestingly, while the inflammatory markers were suppressed, oxamic acid treatment did not affect the phagocytic capacity of LRRK2-G2019S microglia (Fig. 6D). However, the levels of the inflammatory cytokine TNF-α were significantly reduced in LRRK2-G2019S microglia than in the untreated microglia (Fig. 6E). These findings confirm that targeting microglial metabolism can alter the course of immune activation, possibly mitigating the toxic effect of LRRK2-G2019S microglia on dopaminergic neurons.
Finally, to determine whether the rescue of metabolic and inflammatory phenotypes in LRRK2-G2019S microglia has a neuroprotective effect, we assessed dopaminergic neuron survival in midbrain organoids cocultured with microglia and treated with or without oxamic acid. Compared with organoids cultured with healthy microglia, midbrain organoids cultured with LRRK2-G2019S microglia showed significant dopaminergic neuron loss (Fig. 6F). However, treatment with oxamic acid substantially increased the number of dopaminergic neurons in midbrain organoids cultured with LRRK2-G2019S microglia, ameliorating the detrimental impact of LRRK2-G2019S microglia. These data indicate that targeting microglia metabolism in LRRK2-G2019S microglia not only reduces their inflammatory phenotype but also prevents neurotoxicity, demonstrating the potential of immunometabolic approaches that may slow neurodegeneration, pending further validation.
Comments (0)